
Working with staff of Florida Wildflower Foundation, we’ve planned a 6-part Webinar series on the botany of our native wildflowers. In this 4th presentation, discussion moves to plant families – how they are delimited, how many families of plants we encounter in Florida habitats, how a person can determine in which family a plant is likely classified, and how to turn that information into useful knowledge about the identity and nature of a plant.
Skip the Webinar Presentation and go directly to Family Descriptions: Composites (Asteraceae); Euphorbs (Euphorbiaceae); Grasses (Poaceae); Evening Primroses (Onagraceae); Legumes (Fabaceae); Milkweeds & Dogbanes (Apocynaceae); Mints (Lamiaceae); Morning Glories (Convolvulaceae); Orchids (Orchidaceae); Sedges (Cyperaceae);
General discussion begins by regarding a plant “Family” as a branch in the “Tree of Life”, terms that require their own special definition. A “tree of life” portrays evolutionary relationships, showing how we imagine different kinds of life have evolved over time (millions, even hundreds of millions of years), giving rise to the species we encounter today. Examining the branching structure, we discover connectivity that links all living creatures to a common ancestry. Over the eons, the tree has morphed with emergence of new branches and loss of antiquated forms through extinction, the evidence coming from morphological comparisons and molecular study of extant life forms as well as examination of fallen branches we call fossils.

In the living tips of today’s tree, smaller branches represent more tightly-related similar clusters of species which are understood as Genera. Groups of clearly-related Genera can be isolated as a larger branch interpreted as a Family. The biological term “family” then takes the broadest meaning, not limited to the narrow lineages of one human family versus another, like the Hatfields and the McCoys. Rather, a plant Family can be more likened to the “family of man” anthropologists call the Hominidae, the Great Apes, a clearly defined evolutionary clade (branch) that includes humans (Homo), as well as Gorillas (Gorilla), Orangutans (Pongo), and Chimpanzees (Pan).


Biologists have imagined differing trees-of-life. Some trees attempt to represent all of creation, from bacteria to fungi, insects, animals, and plants.

Other diagrams focus on major groups, such as the Plant Kingdom (mosses, ferns, conifers, and flowering plants). As seen in the illustration above, well before Darwin’s Origin of Species was published, and in the midst of Lamarckian discussion, Frenchman Augier taught botany from a diagram that outlined thought current in 1801. Paired with Augier’s depiction, we see a branching diagram from Darwin’s notes and on the right, an elegant 1866 portrayal by Haeckel. Below, a yet earlier representation is taken from Jussieu’s seminal Genera Plantarum… (1789), in which you see an outline of his thoughts concerning flowering plants, a typographic portrayal tantamount to a cladistic tree.

As shown in the slide below, in our current world (2025), a “tree of life” for all life could be more a illustrative conception on the left (similar to Haeckel’s portrayal), or the condensed graphic (similar to Darwin’s sketch) we associate with “cladistic” analyses.


Focusing on flowering plants (Angiosperms), botanists today would draw a “tree” with about 416 living branches labelled as families, organized into stouter limbs called Orders. Some branches are nearly barren, with a single tip (species), while one massively complex branch would be labelled Asteraceae (the Composites), with over 30,000 tips (species) formed on hundreds of secondary branches considered genera. Below we have a generalized contemporary construct available from USBG (the United States Botanic Garden).

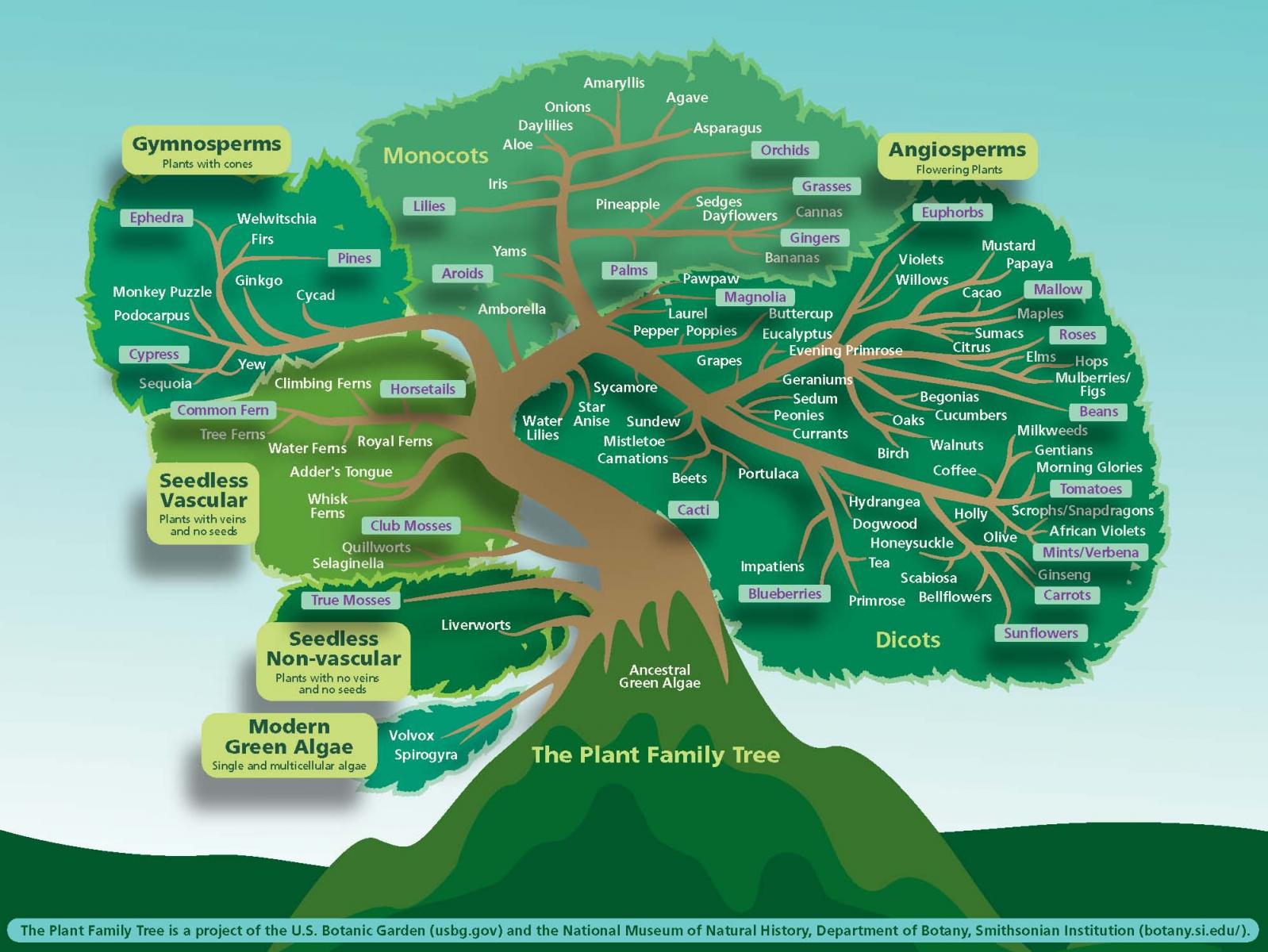{kind=link}
More technical graphics are shown in the following slide. The cladogram to the left graphs Orders (groupings of plant families) as specialists imagine their relationships. The starburst to the right shows plant families radiating (evolving) relative to one another, with colored segments estimating the age of each family.

There’s good knowledge in examining these greater heirarchies. Just as related species are clustered as a Genus (a branchlet on the tree of life), and related genera are envisioned as a single branch, a Family, we imagine families as larger branches, or clades termed Orders, reflecting current thoughts as to the evolutionary ancestry of the plants. At each level, we have the opportunity to examine general characteristics that hold the grouping together, summarizing what is known. More inclusive and formal levels organize Orders into Classes, and Classes into Phyla (or Divisions), and any of the formal categories might be divided into sub-groups, so there’s no shortage of heirarchies that have been proposed. But, as famed botanist Harold Bold told me, “nature mocks at human categories.”

That mocking is apparent from the various depictions of a tree of life, in that relative size and diversity of families is just too much information to convey in a simple graph. By current definition, the five most species-rich plant families (Asteraceae, Orchidaceae, Fabaceae, Rubiaceae, and Poaceae) account for nearly a third of flowering plant species, but diagrams can’t allot so much space to five families when there are more than 400 to showcase. This representational disparity reminds us there’s neither method nor logic for standardizing the definition of species, genera, or families across the plant kingdom. The great diversity of Orchid species (over 20,000 species) is based on pollination biologies completely different from mechanisms that isolate one species of Composite (at over 30,000 species) from another; there’s no simple way to make clear comparisons. So we just ignore this little incongruity.
The Orchids and Composites are not alone; Grasses and Legumes constitute two other very natural groups showing remarkable diversity that would make the tree of life lopsided, with a few massive branches and many that are barely populated. The poorly-represented branches would include 115 plant families that are monogeneric (including a single genus only), with 54 of those represented by only one or two species (Christenhusz and Byng, 2016). Twenty-nine plant families are monotypic (having a single species). Evolution and selection have not been egalitarian and botanists are compelled to reduce every circumstance to a string or graph of words on a two-dimensional platform. Oddly, a realistic rendition of the popular tree of life would seem un-natural.
Restricting a graphic tree to our native flora, we find similar disparities, with several monogeneric families represented – Altingiaceae, Aqufoliaceae, Bataceae, Cannaceae, Ceratophyllaceae, Cleomaceae, Hydroleaceae, Mayacaceae, Nelumbonaceae, Platanaceae, Portulacaceae, Ruppiaceae, and Smilacaceae. Moreover, half of the plant families listed for our native flora are represented by five or less species. And at the other extreme, as is true globally, a few families show great diversity, with hundreds of species – Asteraceae, Poaceae, Fabaceae, Orchidaceae, and Cyperaceae. This conjures a range of challenges for the citizen scientist. Becoming familiar with just ten plant families gives some access to information on half of the native plants, but the remaining half are distributed among 170 families.

How to choose which families we should study first? We begin by examining those with greatest diversity.

Getting Families in Order
Before diving into selecting families for study, there’s a precaution to consider. The parameters of most families are well-established, but over the past three decades, systematists have introduced a wider range of characteristics for consideration (such as molecular/genetic studies), employed particularly in studying family groupings and relationships that haven’t been so clear-cut. That has meant the higher levels for systematizing (organizing) plants have been in considerable flux. The limits of families have been altered, as well as how those families might be grouped to construct Orders and Classes (which would also need to reflect evolutionary origins.) Over the past three decades, ownership of that discussion centered on the Angiosperm Phylogeny Group (AGP), a consortium of taxonomists who consulted each other in order to reach agreement on the meaning of evidence regarding the evolution and systemization of plants. (It isn’t clear those discussions and updates continue.)
Existing formal structures can limit dialogue, with knowledge easily silo’d, thus the AGP promoted a more flexible approach, allowing new information to challenge previous thoughts, promoting ongoing debate that avoids formal structure. Results (now widely accepted) suggest flowering plants can be imagined in three significant assemblages: 1. a cluster of the more ancient plant groups (wetland and woody plants, focusing on the ANA grade (plants in the Amborales, Nymphaeales, and Austrobailieyales) and the Magnolids (Magnolias,Laurels, Pipers, etc.) that gave rise to two better-defined groups, which are 2. the Monocots; and 3. the Eudicots (a tighter assemblage of related dicots).
Following that interpretation, the consortium issued summaries (in publications that are behind paywalls.) You can access a basic summary of the most recent version (2016) though Wikipedia. Moreover, Peter Stevens (Missouri Botanic Garden) maintained discussion of APG issues on a website he updated regularly (until about 4 years ago). Though fluid, it makes sense to organize our discussion of Families based on the APG framework, which parallels the presentation of families in the multi-volume Flora of Florida. (The following link will take you to a PDF Index to families treated in the multi-volume set: Flora of Florida Index)

In the APG framework, we see Florida native Angiosperms (flowering plants) fit into about 180 Families, categorized in 48 Orders that I’ve listed in several major groupings reflecting the following current (but shifting) outline of evolutionary relationships:
- BASAL Angiosperms – Orders (2) in our flora: Austrobaileyales (Schisandraceae) and Nymphaeales (Cabombaceae and Nymphaeaceae)
- Core Angiosperms, MONOCOTS – Orders (5) in our flora: Alismatales (Alismataceae, Araceae, Cymodoceaceae, Hydrocharitaceae, Juncaginaceae, Potamogetonaceae incl. Zannichelliaceae, Ruppiaceae, Tofieldiaceae); Asparagales (Amaryllidaceae incl. Alliaceae, Agavaceae, & Ruscaceae, Asparagaceae, Hypoxidaceae, Iridaceae, Orchidaceae); Dioscoreales (Dioscoreaceae, Burmanniaceae, Nartheciaceae); Liliales (Colchicaceae, Liliaceae, Melanthiaceae incl. Trilliaceae, Smilacaceae); Pandanales (Stemoniaceae)
- Core Angiosperms, Monocots, COMMELINIDS – Orders (5) in our flora: Arecales (Arecaceae); Ceratophyllales (Ceratophyllaceae); Commelinales (Commelinaceae, Haemadoraceae, Pontederiaceae); Poales (Bromeliaceae, Cyperaceae, Eriocaulaceae, Juncaceae, Mayacaceae, Poaceae, Typhaceae, Xyridiaceae); Zingiberales (Cannaceae, Marantaceae)
- Core Angiosperms, MAGNOLIDS – Orders (4) in our flora: Canellales (Canellaceae); Laurales (Calycanthaceae, Lauraceae); Magnoliales (Annonaceae, Magnoliaceae); Piperales (Aristolochiaceae, Piperaceae, Saururaceae)
- Core Angiosperms, Eudicots – Orders (3) in our flora: Buxales (Buxaceae); Proteales (Nelumbonaceae, Platanaceae); Ranunculales (Berberidaceae, Menispermaceae, Papaveraceae, Ranunculaceae)
- Core Angiosperms, Eudicots, Core Eudicots, Pentapetalae, SUPERROSIDS & VITALES – Orders (2) in our flora: Saxifragales (Altingiaceae, Grossulariaceae, Haloragaceae, Hamamelidaceae, Iteaceae, Penthoraceae); Vitales (Vitaceae)
- Core Angiosperms, Eudicots, Core Eudicots, Pentapetalae, Superrosids, Eurosids, FABIDS & N-FIXING+ FABIDS – Basal Fabid Order (1) in our flora: Zygophyllales (Krameriaceae, Zygophyllaceae); Core Fabids (N-Fixing+ Fabid Clade Ordersa0 in our flora (4): Cucurbitales (Cucurbitaceae); Fabales (Fabaceae, Polygalaceae, Surianaceae): Fagales (Betulaceae, Fagaceae, Juglandaceae, Myricaceae): Rosales (Cannabaceae, Moraceae, Rhamnaceae, Rosaceae, Ulmaceae, Urticaceae)
- Core Angiosperms, Eudicots, Core Eudicots, Pentapetalae, Superrosides, Eurosids, COM-CLADE – Orders (3) in our flora: Celastrales (Celastraceae incl. Parnassiaceae); Malpighiales (Chrysobalanaceae, Clusiaceae, Euphorbiaceae, Linaceae, Malpighiaceae, Passifloraceae incl. Turneraceae, Phyllanthaceae, Putranjavaceae, Rhizophoraceae, Salicaceae, Violaceae); Oxalidales (Oxalidaceae)
- Core Angiosperms, Eudicots, Core Eudicots, Pentapetalae, Superrosides, Eurosids, MALVIDS – Orders (8) in our flora: Brassicales (Bataceae, Brassicaceae, Capparaceae, Caricaceae, Cleomaceae); Crossosomatales (Staphyleaceae); Geraniales (Geraniaceae); Malvales (Cistaceae, Malvaceae, Thymeliaceae); Myrtales (Combretaceae, Lythraceae, Melastomataceae, Myrtaceae, Onagraceae); Picramniales (Picramniaceae); Santalales (Olacaceae incl. Ximeniaceae, Santalaceae incl. Viscaceae, Schoepfiaceae); Sapindales (Anacardiaceae, Burseraceae, Meliaceae, Rutaceae, Sapindaceae, Simaroubaceae)
- Core Angiosperms, Eudicots, Core Eudicots, Pentapetalae, Superasterids: Caryophyllales (Cactaceae, Caryophyllaceae, Droseraceae, Nyctaginaceae, Petiveriaceae, Phytolaccaceae, Plumbaginaceae, Polygonaceae, Portulacaceae)
- Core Angiosperms, Eudicots, Core Eudicots, Pentapetalae, Superasterids, ASTERIDS – Orders (2) in our flora: Cornales (Cornaceae, Hydrangeaceae, Loasaceae, Nyssaceae); Ericales (Balsaminaceae, Clethraceae, Cyrillaceae, Ebenaceae, Ericaceae, Polemoniaceae, Primulaceae incl. Myrsinaceae, Samolaceae & Theophrastaceae, Sapotaceae, Sarraceniaceae, Styracaceae, Symplocaceae, Theaceae)
- Core Angiosperms, Eudicots, Core Eudicots, Pentapetalae, Superasterids, Asterids, LAMIIDS – Orders (4) in our flora: Boraginales (Boraginaceae); Gentianales (Apocynaceae, Gelsemiaceae, Gentianaceae, Loganiaceae, Rubiaceae); Lamiales (Acanthaceae, Bignoniaceae, Lamiaceae, Lentibulariaceae, Linderniaceae, Oleaceae, Orobanchaceae, Phrymaceae, Plantaginaceae, Scrophulariaceae, Tetrachondraceae, Verbenaceae); Solanales (Convolvulaceae, Hydroleaceae, Solanaceae)
- Core Angiosperms, Eudicots, Core Eudicots, Pentapetalae, Superasterids, Asterids, CAMPANULIDS – Orders (4) in our flora: Apiales (Apiaceae, Araliaceae, Pittosporaceae); Aquifoliales (Aquifoliaceae); Asterales (Asteraceae, Campanulaceae, Menyanthaceae); Dipsacales (Caprifoliaceae, Viburnaceae incl. Adoxaceae)
After reviewing the range of plant families in Florida Wildflowers as compared to other bases, we will return to the APG platform.
Families in our Native Flora showing the greatest diversity:
The slide below compares plant families in the Florida flora based on which have the most “wildflowers” as compared to “trees & shrubs” as compared to “exotics” (non-native plants). Cutting to the chase, as we examine diversity, it becomes apparent that most of our “Wildflowers” are classified in Families that fall into nine Orders: Poales, Asparagales, Fabales, Malpighiales, Gentianales, Solanales, Lamiales, Apiales, and Asterales. Except for Legumes (in the Fabales) and Rubiacs (in the Gentianales), most of our trees and woody shrubs show up in other orders: Fagales, Ericales, Rosales, Magnoliales, and Laurales. Distinctions are not so clearcut, but the numbers suggest two heartening implications: 1) Gaining some understanding of just 15 Orders of flowering plants opens the door to appreciating the evolutionary histories of well over half of out native flora; 2) The remaining plant groups are smaller families, which pretty much boil down to a few handfuls of Genera. That makes things a lot simpler, right?

The next slide is the same, except using the botanical family names.

The following slides color-code the data given above. In the first we compare the ten most species-rich native plant families to the the top ten families of exotic plants. Colored text indicates which families are in both lists.

Breaking down native plants as Wildflowers vs. Trees & Shrubs, we see Fabaceae (Legumes) as the only family with strong representation among all three groups. The diversity of Legumes in the exotic flora likely relates to their root adaptations for hosting nitrogen-fixing bacteria, which would provide some advantage in nutrient-deficient sandy soils. Both Grasses and Legumes would have been introduced as forage, which may relate to their abundances as escaped exotics.

Comparing the interpretation of “Wildflower” as herbaceous and suffrutescent shrubs to plants treated in Roger Hammer’s Guide to Florida Wildflowers, we see substantial overlap, with the exception of Grasses, Sedges, and Reeds – all of which have reduced (therefore not showy) flowers. The orange-colored text compares the twenty most species-rich wildflower families with families most represented in Hammer’s Guide. (For a complete list of families and species he includes, follow this link to the Hammer Guide to Florida Wildflowers.)

The final slide shows the top ten families in number of species for Florida native plants, as compared to Exotics, as compared to the World flora. Florida natives mirror the world flora in seven families, with a relative lower abundance of Rosaceae, Solanaceae, and Brassicaceae, and greater representation of Euphorbiaceae, Convolvulaceae, and Apocynaceae. Differences suggest Florida’s warm temperate to subtropical climate may not favor Rosaceae and Brassicaceae. The relative absence of native Solanaceae would seem to reflect an impoverished tropical flora in our peninsular state as compared to the intact tropical flora found in great landmass of South and Central America, which boasts considerable diversity of Solanaceae (think about the impact of Tomatoes, Potatoes, and Peppers on world cuisines).

NOTE: If you wonder where I’ve accessed numbers compiled in this document, I’m using an extraordinary resource freely available to us all. Visit the ISP Florida Plant Atlas website, where you can examine data concerning any given Plant Family. We’ll take Lamiaceae as an example. Beginning at the ISB home page, there are two simple ways to construct comparisons. In the search bar, set parameters for “Family” and search for Lamiaceae, you’ll get an astonishing spreadsheet of all taxa in the flora, with many fields, such as Native, Endemic, Status, etc. It’s a great summary. Moreover, you can download that spreadsheet to manipulate. For a completely different comparison, hone in on a single species, and move to the page for that plant. There you’ll see a hierarchy that includes Family and Genus. Click on Family and up pops a spreadsheet showing all Genera as well as the number of taxa. Below that spreadsheet is a key to the Genera. You can copy the cells in that spreadsheet and paste them into Excel or Numbers to manipulate. Below are screenshots of the two Lamiaceae spreadsheets I edited and color-coded:



If pure numbers rule, then it would seem ten groups that demand in-depth study are the three larger Monocot families (Poaceae, Cyperaceae, and Orchidaceae) and seven Dicot families (Asteraceae, Fabaceae, Lamiaceae, Euphorbiaceae, Convolvulaceae, Apocynaceae, and Onagraceae). Generally, these are reasonably natural families, defined by characteristics that make for easy identification. That would seem one logical foundation for exploring characteristics most useful in grouping and identifying plants.
How and where to begin becomes the challenge. If we could have individual conversation, I’d prefer to begin where you are, personally. I’d want to know what you’ve already conquered. That’s not in the cards, so we must go on an assumption anyone taking time to attend or review a presentation on native wildflowers likely already has knowledge of wildflowers. The task then moves to bringing structure and depth to a generalized, basic foundation by exploring well-known and iconic plants, weaving the larger fabric around those keystones. That means, as the slide below says: “You already know many” Plant Families

Thankfully, we have an edge – some Good News. Many of our native Wildflowers fall out in well-known and easily recognized groups (Genera) that characterize very straightforward Families.

The slides below introduces one way to simplify the task of identifying plant Families in field and garden – start with what you know and what you won’t miss by not knowing. In the Webinar, I presented categories of familiarity; the next many slides document that discussion. Afterward, the text presents a more thorough reckoning, in which the families mentioned during the Webinar, as well as others, are described and discussed within the APG framework.

The slide below gives examples of One-Off Plant Families, those limited in numbers and distribution in the native flora. I included Geranium since the two native species are so clearly recognizable and the Family has little other presence in the natural landscape.

The next few slides remind us that many Families are characterized by one or two well-known genera. If you know one or a few of the plants, you have the entire Family under control.









Two Families – the Orcihds and the Composites – are presented as both large (in number of native species) as well as easily recognized.












In the Webinar, I cast three Families as simply too big to ignore, pairing the Grasses and Sedges, then following with the Legumes.









The slide below makes the point that Legumes are better identified by fruit (and foliage) than might be the case for flowers. With experience, the flowers fall into place, but at the outset, there are many distinct forms to learn. The red Callicarpa is a head of flowers, each of which has showy stamens and reduced petals. To the right, the extravagant Bauhinia flower doesn’t immediately suggest close relationship to that of a typical Pea.


Wildflowers are significant members of many other Families; five of the most prominent are showcased below (and described in greater detail in the systematic presentation that follows).

Thsse additional Families – the Mints, the Dogbanes (Apocynaceae), Evening Primroses (Onagraceae), Evening Primroses (Convolvulaceae), and Euphorbs (Euphorbiaceae) – are both common and easily identified.









Finally, I present the Plantaginaceae as Annoying, in that it seems to have become the dustbin for plants that were scrubbed from other Family associations. Odd-seeming, morphologically, to group Plantago with Scoparia, Digitalis, and Callitriche. It simply does not seem to be a natural grouping.

The presentation ended with Acknowledgements. Following this slide you will find a systematic treatment of Florida Wildflowers, after which there are References and Resource notes.

Flowering Plants: Basal Angiosperms, Magnolids, Monocots, and True Dicots (Eudicots)
Plants that seem to reflect the most ancient nature of Angiosperms (the Flowering Plants) are characterized as Basal, a broadly diverse group of plants that lack characteristics botanists consider more evolutionarily advanced, features we see in the more “derived” Monocots and “True” Dicots.
Basal Angiosperms
Among Basal Angiosperms we find plants botanists have short-handed as ANA-grade (or ANITA-grade), an assembly showing vegetative and floral characters botanists associate with earlier-evolved flowering plants – sometimes called “paleoherbs” or “paleodicots.” These living relicts include the singular New Caledonian Amborella, as well as common Florida plants in the Nymphaeales and Austrobaileyales (thus the acronym ANA). When Illicium was recognized in its own family (Illiciaceae), at a time the basal group was just being defined, you may have heard botanists using the term ANITA-grade (the “I” representing Illiciaceae).




Among the Nymphaeales, the ISB Atlas still recognizes Cabombaceae (with two Natives, i.e. Cabomba and Brasenia and one Exotic Cabomba species) as separate from Nymphaeaceae, which includes the native Nuphar advena (3 varieties) and five native Nymphaea (plus two Exotic species).

The plant we call Spatterdock, Nuphar, though often encountered in the Panhandle, is ever-present in the Everglades. Below is a photo I made of our Nuphar advena, followed by a labelled dissection of a Nuphar variegata, published in the Flora of Newfoundland and Labrador website.


Magnolids


Right there with the muddled ANA-grade plants is a better-defined branch (read “monophyletic”) nicely present in our North Temperate flora, the Magnolids. Most of these are woody (Magnolias, Laurels, Annonas), which casts them outside the strict definition of Wildflowers, but Hammer includes Asimina in his book. And native Piperales include the herbaceous Peperomia, Pipeworts (Aristolochia, Asarum), and the wetland Lizardtails (Saururus).

Here’s an outline to the Orders of Magnolids and their representation in our flora (annotated with numbers of Genera/Species):
- Canellales: Canellaceae (Native 1/1 Canella)
- Laurales Calycanthaceae (Native 1/1 Calycanthus), Lauraceae (Native 7/10 including Persea, Sassafras, Lindera; Exotic 3/4 including Avocado, Persea americana, and the invasive evergreen, Camphora officinarum (Cinnamomum camphora))
- Magnoliales Annonaceae (Native 2/12 Annona, Asimina includes 6 hybrids; Exotic 3/3 ), Magnoliaceae (Native 2/7, Magnolia, Liriodendron)
- Piperales Aristolochiaceae (Native 2/5 Aristolochia, Asarum; Exotic 1/5 Aristolochia), Piperaceae (Native 1/6 Peperomia; Exotic 2/6 Piper, Perperomia), Saururaceae (Native 1/1 Saururus; Exotic 1/1 Houttynia)

The Inevitable Monocots and Dicots:
Dispensing with the two Basal groups leaves a pair of distinct and major branches: the Monocots and the Eudicots (True Dicots) – familiar territory. These are the two classes of flowering plants I first learned to discriminate – the curious Monocotyledons, given their singular, tubular-leaf growth, parallel veins, scattered vascular bundles, and 3-merous approach to flower parts – all of that as contrasted with the textbook-normal Dicotyledons, with seed sprouting an initial pair of seed leaves (cotyledons) followed by a more regular leaves along stems capable of making conventional woody trees (like oaks), everyday shrubs (like boxwood), regular vines (like Vitis or Campsis), and ordinary herbs (like mustard), and producing flowers with green sepals.
The Monocots I first knew botanically remain a natural assemblage. We think of them as constituting two major clades (botanists share a “dichotomy-bias”, a compunction to break every complex assortment into two contrasting assemblages): the “Lily” sorts and the “Commelina” branch. To me, the Lilies are the horticulturally-precious (Trillium, Crinum, Aroids, Orchids), versus the other branch, the Commelinids, which strike me as more industrial (Gingers, Palms, Grasses, and Sedges).
As will be discussed later, the APG also groups Eudicots with a basal cluster from which arise two main branches (clades), the Rosids and the Asterids. That makes for three groups of familiar dicots: 1. the basal Lotus and Clematis families, surmounted by 2. the Rose clade, producing polypetalous flowers (with observable petals that are typically free), including Roses, Legumes, Mallows, & Euphorbs, and 3. the Asterids, plants with tubular corollas, such as Mints, Morning Glories, and Daisies. We will get to this assortment of Dicots.
For the moment, however, there’s plenty to consider in the world of Monocots, only a fraction of which would be trees or woody plants. That means the great bulk of Florida Monoc0ts are bona fide Wildflowers or Wilded-flowers (non-native ruderals and weeds that populate our waysides).
MONOCOTS
Among native Wildflowers, five Orders (at least 16 Families) are represented in the Core Monocots, and another five (encompassing 15 Families) loosely grouped as Commelinids. This disposition has been accepted generally, following more than a century of lingering turmoil surrounding delineation of certain included Families and their assignment to Orders.
That’s not to say there was general disagreement. Some Monocot groupings have never been challenged; Palms (Arecales) have long been regarded as an indisputably natural branch in the tree of life; Jussieu recognized the Palm Family (he called it an Order, the Palmae) in his 1789 Genera Plantarum, the publication botanists accept as the foundation for classifying plants. Grasses and Gingers haven’t been challenged.


But other classifications have proven difficult, most particularly relationships among the plants we call “petaloid monocots” – those with showy, similarly-colored tepals such as Lilium, Trillium, Hemerocallis, Gloriosa, Agave, Yucca, Aloe, Amaryllis, Hippeastrum, Crinum, and Hymenocallis – a host of horticulturally-important plants. Limits of Liliales and Asparagales, as well as definition of Families in those Orders have shifted radically with the advent of plant “phylogenetic” studies (the contemporary use of genetic analysis combined with traditional descriptive information to map out evolutionary relationships among plants) over the past three decades.
The dust having settled a bit, several small families traditionally considered Liliales are now thought more aligned to Asparagales, an order that has come to dominate the petaloid monocot landscape. In the outline below, you’ll see the expansive Asparagales, and the consolidated Poales to be well-represented among Florida’s Wildflowers. Here’s a reminder of the current systematic state and representation in our flora, beginning with the Lily Orders (Genera/Species):
- Acorales: (Not Represented)
- Alsimatales: Alismataceae (Native 3/12 Echinodorus, Helanthium, Sagittaria; Exotic 2/2 Echinodorus, Sagittaria), Araceae including Limnaceae (Native 8/19 Arisaema, Lemna, Orontium, Peltandra, Pistia, Spirodela, Wolffia, Wolffiela Exotic 15/18 Aglonema, Alocasia, Anthurium, Colocasia, Cryptocoryne, Dieffenbachia, Epipremnum, Gonatopus, Landoltia, Lasia, Monstera, Philodendron, Syngonium, Wolffia, Xanthosoma), Hydrocharitaceae (Native 6/11 Elodea, Halophila, Limnobium, Najas, Thalassia, Vallisneria; Exotic 4/4 Egeria, Hydrilla, Najas, Ottelia), Potamogetonaceae (Native 2/10 Potamogeton, Stuckenia; Exotic 1/1 Potamogeton), Ruppiaceae (Native 1/1 Ruppia), Tofieldiaceae (Native 3/3 Pleea, Harperocallis, Tofieldia (Triantha))
- Dioscoreales: Dioscoreaceae (Native 1/2 Dioscorea; Exotic 1/5 Dioscorea, including the invasive Air-Potato, Dioscorea bulbifera), Burmanniaceae (Native 2/4, Aptera, Burmannia), Nartheciaceae (Native 2/6 Aletris, Lophiola)
- Pandanales: Stemonaceae (Native 1/1 Croomia)
- Liliales: Liliaceae (Native 3/7 Erythronium, Lilium, Mediola; Exotic 1/1 Lilium), Colchicaceae (Native 1/3 Uvularia; Exotic 1/1 Gloriosa), Smilacaceae (Native 1/12 Smilax), Melanthiaceae (Native 6/13 Amianthium, Chamaelirium, Schoenocaulon, Stenanthium, Trillium, Veratrum, Zigadenus)
- Asparagales: Asparagaceae (Exotic 1/3 Asparagus), Ruscaceae (Native 3/4 Maianthemum, Nolina, Polygonatum; Exotic 4/7 Aspidistra, Dracaena, Liriope, Ophiopogon), Agavaceae (Native 3/7 Agave, Schoenolirion, Yucca; Exotic 3/8 Agave, Chlorophytum, Fucraea), Hyacinthaceae (Exotic 1/1 Muscari), Alliaceae (Native 2/4 Allium, Nothoscordum; Exotic 4/4 Allium, Navalia, Ipheion, Nothoscordum), Amaryllidaceae (Native 3/14 Crinum, Hymenocallis, Zephyranthes; Exotic 6/17 Crinum, Galanthus, Leucojum, Lycoris, Narcissus, Zephyranthes), Asphodeleaceae (Exotic 2/4 Aloe with horticultural hybrids, Bulbine), Iridaceae (Native 4/14 Calydorea, Iris, Nemastylis, Sisyrinchium; Exotic 8/12 Iris, Sisyrinchium, Crocosmia, Dietes, Freesia, Gladiolus, Herbertia, Trimezia, including horticultural hybrids), Hypoxidaceae (Native 1/5 Hypoxis; Exotic 1/1 Curgulio), Orchidaceae (Native 50/114 Basiphyllaea, Beloglottis, Bletia, Brassia, Bulbophyllum, Calopogon, Campylocentrum, Cleistesiopsis, Corallorhiza, Cranichis, Cyclopogon, Cyrtopodium, Dendrophylax, Eltroplectris, Encyclia, Epidendrum, Eulophia, Galeandra, Goodyera, Govenia, Habenaria, Hexalectris, Ionopsis, Isotria, Lepanthopsis, Liparis, Macradenia, Malaxis, Maxillaria, Mesadenus, Neottia, Oncidium, Orthochilus, Pelexia, Platanthera, Platythelys, Ponthieva, Prescottia, Prosthechea, Sacoila, Spiranthes, Stelis, Tipularia, Tolunia, Trichocentrum, Triphora, Tropidia, Vanilla; Exotic 11/13 Bletia, Bletilla, Cyrtopodium, Encyclia, Epidendrum, Eulophia, Laelia, Oeceoclades, Phaius, Spathoglottis, Vanilla, Zeuxine)
And then, the Commelinoids:
- Arecales: Arecaceae (Palms, Native 9/11 Acoelorraphe, Coccothrinax, Leucothrinax, Pseudophoenix, Rhapidophyllum, Roystonea, Sabal, Serenoa, Thrinax; Exotic 14/16 Acrocomia, Adonidia, Butia, Caryota, Chamaedorea, Cocos, Dypsis, Elaeis, Livistona, Phoenix, Ptychosperma, Saribus, Syagrus, Washingtonia)
- Commelinales: Commelinaceae (Dayflowers, Native 3/10 Callisia, Commelina, Tradescantia; Exotic 5/18) Callisia, Commelina, Gibasis, Murdannia, Tradescantia, Haemodoraceae (Redroots, Native 1/1 Lachnanthes), Pontederiaceae (Pickerelweeds & Water Hyacinth, Native 2/3 Heteranthera, Pontederia; Exotic 2/4 Eichornia)
- Poales: Bromeliaceae (Bromeliads, Native 2/17 Catopsis, Guzmania, Tillandsia; Exotic 4/4 Billbergia, Bromelia, Dyckia, Tillandsia), Typhaceae Native 2/3 Sparganium, Typha), Eriocaulaceae (Native 3/12 Eriocaulon, Lachnocaulon, Syngonanthus), Xyridaceae, (Native 1/25 Xyris), Juncaceae (Reeds, Native 2/24 Juncus, Luzula), Cyperaceae (Sedges, Native 17/244 Abilgaardia, Bolboschoenus, Bulbostylis, Carex, Cladium, Cyperus, Dulichium, Eleocharis, Fimbristylis, Fuirena, Isolepis, Rhynchospora, Schoenoplectiella, Schoenoplectus, Schoenus, Scirpus, Scleria; Exotic 6/42 Bulbostylis, Cyperus, Eleocharis, Fimbristylis, Fiurena, Scleria), Poaceae (Grasses, Native 72/265; Exotic 79/196 – Genera with Natives Only: Alopecurus, Amphicarpum, Anthenantia, Arundinaria, Axonopus, Brachyelytrum, Chasmanthium, Coelorachis, Coleataenia, Ctenium, Danthonia, Dichanthelium, Diplachne, Disakisperma, Distichlis, Elionurus, Elymus, Gymnopogon, Leersia, Leptochloa, Limnodea, Muhlenbergia, Paspalidium, Phanopyrum, Pharus, Piptochaetium, Schizachyrium, Sorghastrum, Spartina, Sphenopholis, Stapfochloa, Stenotaphrum, Tridens, Triplasis, Tripsacum, Uniola, Zizania, Zizaniopsis; Genera with both Native and Exotic Species: Agrostis, Andropogon (one exotic), Aristida (one exotic), Bothriocloa, Bouteloua, Bromus, Cenchrus, Chrysopogon, Digitaria, Dinebra, Echinochloa, Eragrostis, Eriochloa, Eustachys, Festuca, Glyceria, Hordeum, Hymenachne, Kellochloa, Lasiacis, Luziola, Oplismenus, Panicum, Paspalum, Phalaris, Phragmites, Poa, Saccharum, Sacciolepis, Setaria, Sporobolus, Steinchisma, Urochloa: Genera with All Exotics: Aira, Alloteropsis, Anthephora, Anthoxanthum, Arthraxon, Arundo, Avena, Bambusa, Briza, Chloris, Cortaderia, Cymbopogon, Cynodon, Dactylis, Dactyloctenium, Desmazeria, Dichanthium, Eleusine, Eremochloa, Hackelochloa, Hemarthria, Heteropogon, Hyparrhenia, Imperata, Koeleria, Lagurus, Lithachne, Lolium, Melinis, Microstegium, Miscanthus, Neyraudiam Oryza, Phleum, Phyllostachys, Pleioblastus, Polypogon, Polytria, Pseudosasa, Rottboellia, Secale, Sorghum, Themeda, Tripidium, Triticum, Zea, Zoysia)
- Zingiberales: Musaceae (Plantains, Exotic 1/3 Musa, includes the hybrid M. ×paradisiaca), Zingiberaceae, (Exotic 6/6 Alpinia, Curcuma, Globba, Hedychium, Kaempferia, Zingiber) Marantaceae (Native 1/1 Thalia; Exotic 1/1 Maranta), Cannaceae, (Native 1/1 Canna; Exotic 1/3 Canna, including a horticultural hybrid), Costaceae (Exotic 1/1 Costus), Heliconiacae (Exotic 1/1 Heliconia), Strelitziaceae (Strelitzia is planted widely, but not reported as introduced to the flora)
We’ll review Wildflower representation in these Monocot Families, examining the Orchids (Asparagales) and two Families in the Poales (the Grasses and the Sedges) in greater detail.
ALISMATALES
Ecologically, Plant Families classified as Alismids trend to the water-dependent. The six families in this Order (Alismataceae, Araceae, Hydrocharitaceae, Potamogetonaceae, Tofieldiaceae, and Ruppiaceae) comprise over eighty species of herbaceous natives and exotics, ranging from aquatic and wetland specialists to denizons of wet prairies and the forest floor to vines and epiphytes, showing a range of floral reduction and adaptations that suit the habitat. The most diverse groups are the aquatic Hydrocharitaceae (interpreted as nine genera, six being native) and the Araceae (twenty-two genera of plants spanning habitats and form, from the miniature floating native Lemna, Wolfia, and Wolfiella, to the North Temperate Arisaema, and the the non-native tropicals, such as Philodendron and Syngonium.)
Within the Alismids, Araceae stand out as distinct, the flowers bearing a single pistil of fused carpels, while flowers in the other families produce separate (free) carpels.






Though Araceae are clearly a distinct element compared to other families included in Alismatales, the Tofieldiaceae stand out also. This is the family in which botanists have grouped North Florida’s Pleea, Tofieldia (Triantha), and the endemic Harperocallis, plants that would seem equally comfortable in the Liliales.


DIOSCOREALES
This Order raised some curious issues as I was chasing down resources and information for this page. The first is simply technological and utilitarian. We’re so comfortably enabled by digital devices that it’s easy to lose track of real information. Searching in POWO to determine more about the nomenclature of Dioscorea, we learn Kew credits the generic name to Plumier, who illustrated a plant he called ‘Dioscorea scandens, foliis tamni, fructu racemoso‘ in his 1703 Nova Plantarum Americanum Genera. In the text, Plumier explains the eponym honors Pedanius Dioscorides, the great early Greek Physician who authored de Materia Medica. By the time Linnaeus published Species Plantarum (1753), in which he accepted Plumier’s genus while adapting the name of his plant to Dioscorea sativa (signifying this is a cultivated plant), nine species were known (of the 600 species recognized today). When I searched Google for ‘Dioscorea scandens Plumier’ however, the AI interface invented different stories, disregarding the Plumier association and at first suggesting this might be a name for Hottentot Bread, South Africa’s D. elephantipes. The real tale, however, is that Linnaeus’s Dioscorea villosa (native to our flora) is recognized as the type for the Genus, and subsumes Dioscorea sativa. Lessons emerge: 1) our native Dioscorea is the basis for the generic and familial names, dating to Plumier’s 1703 publication; 2) even names in Linnaeus’ 1753 Species Plantarum can be subsumed; and 3) don’t take AI generated information as gospel.


The second curious issue, for me, is the seemingly strange bedfellows that emerge in contemporary phylogenetics. You are welcome to read the extensive and content-solid Wikipedia entry on Dioscoreales, but the information provided doesn’t make me any more comfortable with the idea this order includes the Nartheciaceae (Aletris and Lophiola), native wildflowers that seem out of place with Dioscorea and Burmanniaceae.



PANDANALES
Our flora records presence of two families currently assigned to this Order, the Pandanaceae (with one non-native Pandanus) and Stemonaceae, which includes our sole native, the somewhat rare and ever mysterious Croomia.




LILIALES
Until the advent of contemporary phylogenetics, the Liliales was a much broader construct, including most of what has now been assigned to the Asparagales. Today, at least in our regional flora, the Order embraces four Families – Liliaceae, Colchicaceae, Smilacaceae, and Melanthiaceae. Wildflower enthusiasts throughout much of the state will be familiar with the iconic Lilium catesbaei (Liliaceae), and those who venture close to Alabama and Georgia may have encountered one of the Bellworts (Uvularia, Colchicaceae).

Everyone will have tangled with Smilax, a genus that owns its family and blankets the state with a snarl of species (twelve listed) over which I have no confidence. My confustication with Smilax is amplified by their dioecious strategy. Not only are leaves and vine structure variable, but staminate and pistillate flowers are simple, tiny, and borne on different individuals. It’s an important, yet under-characterized group.


Perhaps the most-encountered of the families would be the Melanthiaceae, including the lily-like Amianthium and Stenanthium, our zany Zigadenus, and the misfit but beautifully triptych Trillium. Tell me how these belong together?


ASPARAGALES
If there’s a parallel to “charasmatic megafauna” (Tigers, Zebras, Elephants…), it’s those Petaloid Monocots, many of which are in the Asparagales. Think Crinum americanum and Iris tridentata.


We have nice representation of those two families, with fourteen native species each, but the largest family in the Asparagales would be the Orchids, with over 100 species native to Florida, and over 20,000 worldwide.
Orchidaceae
There’s something special about Orchids that makes each encounter a gift. Perhaps it’s a combination of the allure we attach to the Paphiopetalums, Cattleyas, Cymbidiums, and other large-flowered exotic forms that drove a craze for tropical plants in the 19th century, or the relative rarity of most kinds in the wild (despite the incredible diversity), or their well-known specialized and fascinating pollination systems, or simply the fact that the flowers are spectacular and curious. Consistently surprising, there’s much about Orchids that’s easy and delightful to study.

I’m introducing the family with an exotic Cymbidium in order to clearly describe floral structure. In the presentation slide below you’ll see this plant offers a streamlined example of Orchid flowers, against which we can compare the many kinds of native Orchids one might encounter in Florida.

The basic plan for an Orchid flower is that of the petaloid monocots, with 6 tepals discernable as 3 outer segments (the sepals) and 3 inner segments (the petals). But with Orchids, the three sepals are especially distinct from the three petals. Moreover, the dorsal sepal, which lines up (behind, or above) with the column usually differs from the two lateral sepals. Similarly, one of the 3 petals (the labellum, which is opposite the “column”) is highly modified compared to the pair of lateral petals. The modification relates directly to the pollination system in many formats because the labellum often provides a landing and positioning platform (which would include any calluses or shaped edges), and actually may produce a nectar-rich spur which provides a caloric reward. Usually florid, the labellum might also be reduced and pulled away to accommodate visitation by hovering insects or birds. The labellar structure (as well as that of the other tepals) totally complements the specialized nature of an Orchid’s sexual parts, which form a united mechanism engaging the column. In more recently evolved Orchids, that column is sleek and easily comprehended, housing a stigma-lined cavity surmounted by a single anther. In more ancient types, the column more clearly reflects evolution from plants with three, then two anthers. Because there are so many terrestrial orchids with what’s interpreted as two stamens, some botanists break the group into at least two families. In this description, I’m not following that lead. If it’s a Monocot with pollinia and a defined column, this presentation will consider it an Orchid.
The portion of the column that is clearly the anther is highly derived (specialized) compared to almost any other flower group (except the Asclepiads.) In more advanced Orchid groups (like Cymbidum) it’s simply as an anther cap. With Platanthera, which you can see in the slide below, the anther is split as two separate chambers. But in Platanthera as well as Cymbidium, the pollen forms in masses called pollinia. Those masses are somewhat mealy in Platanthera, though still clinging together (botanists call this “sectile” pollen). In more recently evolved taxa, especially the epiphytic and semi-terrestrials like Cymbidium, the pollinia are solid, even hard and waxy.


That means the pollen load of an orchid might be delivered in one or a very few contacts with a stigma. But do not underestimate the impact; most orchids can generate a thousand or more seed (up to 1-2 million with many of the tropicals) in a single fruit (a capsule) from a single pollination event. That all-or-nothing strategy contrasts with tactics we see in Grasses and other wind-pollinated plants. Orchids can live and reproduce successfully in scattered populations while we find wind-pollination often associated with plants of high local density. And, of course, orchids have a future tethered to particular insects. It’s a small social group, the flower and pollinator. In fact, the “specificity” we observe in orchid pollination systems is the key to understanding barriers (or lack of barriers) between different species. Indeed, orchids are notoriously interfertile, normally readily setting seed with pollen from a related species if an interloping insect visitor (or a human) transfers pollen from one species to the stigma of another.
Of course, it isn’t simply the pollen and pollination mechanisms that define this group of plants. Orchids are rare among seed plants in producing pauperized seed – they produce thousands, even millions of seed that are minuscule, lightweight packets of undifferentiated cells (neither developed embryo nor energy storage). Windborn and free, they are the truest of vagabonds – indigent, unfettered, and rife with possiblity. An orchid seed will only establish successfully if adopted by a fungus that provides nourishment (or is planted in nutrient-rich sterile media by a technician). That gives these plants incredible dispersal capacity coupled with low-survival. But it works. A lot of seed, dispersed widely has ensured success over millions of years.
Considering the particulars involved, Orchids have invested heavily in a lifestyles and strategies that are dependent on intact ecosystems. An Orchid species relies on the “services” of one or few pollinators for reproduction and the presence of the right microhabitat with accommodating fungi for establishment. Every Orchid I encounter is a poignant reminder that like Polar Bears and Penguins, Orchid diversity (estimated at over 25,000 different forms we consider species) faces serious depletion as habitat destruction and climate change progress over this century.
Orchids show up in most areas of the world; those found in more temperate areas being likely terrestrial, and appearing to be more ancient in the evolutionary timeframe. With over 100 species of native Orchids and a climate spanning from tropical to temperate zones, Florida hosts a wide range of types. Some are showy while others defy expectations, being of modest size and appearance, even camouflaged, even achlorophyllous. As long as the right pollinator can locate a flower, all is copacetic.
Useful Terminology for Studying Orchids
- Orchidology – the field of study for an Orchidist
- Corm – a somewhat flattened, enlarged stem of several contracted nodes at or just below ground surface
- Tuberous – fleshy swollen stems are “tubers” – many terrestrial orchids have fleshy, swollen roots resembling tubers, which leads to the term “tuberous” root. The swollen, starchy roots, characteristic of many European terrestrial orchids is the origin of the Greek orchis (which is Greek for “testis”) for these plants
- Pseudobulb – the succulent, united swollen stem-like sheaths of many orchids (typically epiphytic orchids), of few to several nodes, in some species contracted with all blades and scars apical to the pseudobulb, in others with visible internodes showing leaf blades or scars alongside the swelling
- Vellum – a notable spongy epidermal layer characteristic of epiphytic and semiterrestrial orchids, aids in water absorption and indicative of a healthy plant – in orchids, healthy roots are requisite for a healthy plant
- Eye – in epiphytic, pseudobulbous orchids, an axillary bud (or terminal bud in deciduous orchids) that is visibly swollen, promising new growth or flowering
- Conduplicate vs Plicate – leaves that fold along a central vein (conduplicate) versus leave blades that are lightly or strongly corrugated based on several strong veins
- Resupinate – repositioned 180°. Most orchid flowers position the labellum lowermost when the flower is open, but developmentally, the labellum is atop the bud. You’ll see a twist in the ovary (stalk), or perhaps the entire inflorescence with be pendent, orienting the flower such that the labellum is lowermost. Platanthera nivea and some other orchids are non-resupinate – the lip is uppermost in the developed flower
- Dorsal vs Lateral Sepal – Dorsal suggests the top, the ‘adaxial’ surface. In orchids, the sepal subtending the column , and being uppermost in resupinate flowers, is the Dorsal Sepal. It is usually more prominent than the Lateral Sepals. The laterals are the pair of sepals (yes, tepals) on each side of the labellums. The pair of petals flanking the Dorsal Sepal would be the Lateral Petals
- Labellum (lip) – the defining petal of orchids that faces the column
- Callus – a common 3-dimensional surface feature particular to orchid labellums. Often differentially-colored and textured, the callus will have some role in orienting and positioning pollinators.
- Peloric – a term for orchid flowers in which the two lateral petals have taken on the special characteristicds of the labellum, a curious genetic feature of selected hybrid plants
- Spur – the most ommon term for a nectary that shows as a pointed or tubular feature of the labellum
- Nectary – a site for production of a reward (typically nectar) for pollinators, may be a shallow depression but normally a spur. Some orchids bear glands that offer other rewards, such as essential oils, or would be the particular sites of fragrance production
- Orifice – the entrance, opening, to the spur/nectary
- Column – the central feature of an orchid flower, a single structure uniting anther, style, and stigma
- Pollinium – a single serving or orchid pollen, a single structure or parcel, differening from genus to genus
- Pollinarium – the entire pollen apparatus, including any stipe or viscidium, and all pollinaria
- Sectile – describing the loose but coherent pollen in more ancient orchids, grains held together by viscid strands
- Stipe – an elegant and miniscule piece of tissue that serves to attach a pollen load (one or more pollinia) to a viscid surface for attachment to a pollinator. It’s common for the stipe to change in position once attached and removed, making deposition more efficient. The stipe is structurally distict from the non-cellular strands of viscin that form the caudicle, attaching the pollen mass to the stipe
- Viscidium – a structurally-distinct sticky pad that attaches the stipe (pollinarium, pollinium) to a vector
- Rostellum – the tissue between anther and stigma in a column, housing the stipe and viscidium
- Capsule – a simple fruit type that dries and dehisces (splits along sutures) when mature; Orchids have inferior fruit made of three united carpels
- Mycorrhizal – a term uniting myco (Gr. fungus) with rhizi (Gr. root) used to denote presence of a natural association (usually beneficial, thus symbiotic) between fungi and plant roots
poales
Poaceae

We’ll be learning Grasses together, as I’ve spent decades pretending to be a botanist without a real appreciation for these plants (along with the Sedges and Rushes). It’s not an odious task, but a challenge nonetheless. Grasses, so very productive, and so important for sugars and carbohydrates that stoke human diets with calories, are also unheralded, perhaps because they are so very common, or more likely because their wind-pollinated flowers are reduced to the point that we hardly take note of them. Grasses, however, are not to be ignored; they are relentlessly productive, purely sexual. and ultimately fruitful. These plants are all about stems, blades, stamens, stigmas, and grains – no building of massive secondary wood, no time or energy wasted on showy petals and sepals, no kowtowing to insects that require a reward for pollination services.
Hyper-industrial, Grasses remind me of Carl Sandburg’s Chicago
Hog Butcher for the World,
Tool maker, Stacker of Wheat,
Player with Railroads and the Nation's
Freight Handler;
Stormy, husky, brawling,
City of the Big Shoulders:
They tell me you are wicked and I believe them, for I have seen your painted women under the gas lamps luring the farm boys.
And they tell me you are crooked and I answer: yes, it is true I have seen the gunman kill and go free to kill again.
And they tell me you are brutal and my reply is: On the faces of women and children I have seen the marks of wanton hunger.
And having answered so I turn once more to those who sneer at this my city, and I give them back the sneer and say to them:
Come and show me another city with lifted head singing so proud to be alive and coarse and strong and cunning.
Flinging magnetic curses amid the toil of piling job on job, here is a tall bold slugger set vivid against the little soft cities;
Fierce as a dog with tongue lapping for action, cunning as a savage pitted against the wilderness,
Bareheaded,
Shoveling,
Wrecking,
Planning,
Building, breaking, rebuilding,
Under the smoke, dust all over his mouth, laughing with white teeth,
Under the terrible burden of destiny laughing as a young man laughs,
Laughing even as an ignorant fighter laughs who has never lost a battle,
Bragging and laughing that under his wrist is the pulse, and under his ribs the heart of the people,
Laughing!
Laughing the stormy, husky, brawling laughter of Youth, half-naked, sweating, proud to be Hog Butcher, Tool Maker, Stacker of Wheat, Player with Railroads and Freight Handler to the Nation.
Seemingly a bit hyperbolic? Not really! Grasses produce! Warm-season Grasses (specifically the PACMAD Clade) along with many Sedges, ill-content with the slower pace of everyday photosynthesis adopted the industrial C4 approach, aggressively capturing and fixing carbon dioxide for energy-hoarding sugars. A cornfield or sugarcane brake of grasses, given nutrients, water, and relentless sunshine will be among the most productive biological systems on Earth. That doesn’t diminish the importance of their cool-season C3 cousins like Wheat, Rice, and Oats, which rival Corn as our most important grains. Grasses are aggressive, hell-bent on growing;
Why the pressing hurry? Why so productive? Well grasses cover a lot of territory. I’ve seen estimates suggesting that with overwhelming presence in savannas, steppes, llanos, and prairies, grasses dominate as much as 40% of Earth’s landmass. They don’t, of course, grow fast to cover territory, rather they cover territory because they’ve evolved the capacity to grow fast, along with architecture that allows them to conquer square footage in many different habitats.
It helps that Grasses are Monocots. Their parallel leaf veins are a perfect factory design for high photosynthetic productivity. The inherent ability to roll leaves when water is restricted is a useful tactic to combat water loss. Their aerial stems (culms) give height and produce flowers, while as Monocots grasses are particularly good at producing multiple leaders that we call stolons (runners) when above ground and rhizomes when burrowing. Perhaps most importantly, Monocots are experts in producing roots at nodes. We call these adventitious roots, which nominally means they show up in unexpected places, but there’s nothing unexpected about rooting nodes in grasses. In fact, ecologists give up on determining what to count as a single grass plant, simply scoring the cluster of upright leaves at each node as a “ramet.” And it isn’t only the ability to generate new growing tips, some grasses even develop intercalary meristems, replacing stem or blade lost to grazing (or the mower).
There is of course a downside to their success. People have introduced non-native grasses to pastures in countries all over the world as crops and forage, where they’ve proven to be expert pioneers. Nearly two hundred species of non-native plants in the Florida flora are Grasses, the most of any plant family.
I headlined this section by stressing the different, almost industrial character of flowering and reproduction. Accustoming ourselves to floral structure in these plants requires learning a new vocabulary and visualizing a different model. Forget petals and sepals as conspicuous elements; think of scales (bracts) creating an armature that houses the stamens and protects the ovaries. Concentrate on an armadillo of scales called the “spikelet“. Knowing and differentiating spikelets is the key to grass taxonomy, the key to identifying the nearly 500 species of native and non-native grasses growing in Florida.
If you want to go full throttle into grasses, get a copy of Agnes Chase’s primer – First Book of Grasses; The Structure of Grasses Explained for Beginners (1922), a near-century-old classic that remains useful as a thorough introduction to the Poaceae. (it’s available for about $10 in Kindle, and at no cost from Smithsonian’s Biodiversity Heritage Library as well as the Internet Archive.) Yes, names have changed, and classification within the family as well, but Chase’s explanation of spikelet types rings true and clear. I’ll be tracing her steps in this summary.

Agnes begins with some rules grasses hold to, not invariably, but generally. Firstly, the blades along the stem are sheathing, the sheathes being open (split) to the point of attachment at a node. At the juncture of blade and sheath is a modest decoration, sometimes a flap of tissue, other times a line of scales; regardless how expressed it may be, this ligule requires attention as a deciding factor in some identifications. Leaves (sheath & blade) are arranged distichously along the stem, which means there are only two ranks. The same will be true for the bracts of most spikelets, which means developing spikelets are basically planar. Secondly, a spikelet typically bears two non-floral basal bracts, the first and second “glumes”, a term derived from glubere, the Latin term for the husk of a grain (derived from a verb that meant “to peel”.)
The glumes are attached to a slender stem called the “rachilla” subtending the remaining spikelet of one or more flowers that may be perfect, imperfect., or even sterile. Each flower produced on a rachilla is encased by its own pair of bracts, the larger (to the outside, abaxial) termed a “lemma” (another term related to husking or peeling) and the inner (adaxial), usually smaller bract called a “palea” (chaff, in Greek). Together, this unit (lemma, palea, & flower) is called a “floret”. See illustrations from Chase, below. Note the text for Fig. 3 refers to the “rudiments of floral envelope”, which references the “lodicules”, bits of tissue that swell to force the lemma and palea open so mature anthers and receptive stigmas are exposed. The lodicules are interpreted as reduced flower parts equivalent to the petals or sepals. But they are uncelebrated and seldom mentioned in general discussion.





In summary, Chase reminds us: “The spikelet is the unit of the inflorescence; the floret is the unit of the spikelet. The spikelet is always simple; that is, the rachilla never branches. The floret is always 1-flowered with never more than one lemma and one palea; the glumes and florets are always alternate, two consecutive ones never being borne one above the other.” Her lesson III, Modifications of the Spikelet, is a spectacular summary of basic structure, from which I take a few pointers. Numbers of florets in spikelets will vary from one to more than twenty. Parts vary in prominence; the palea may be surpressed (undeveloped) in some grasses, but the lemma is always present. The rachilla may be stiff and pointed, but the greatest potential extension comes with lemmas, which may terminate with long bristles called awns. Spikelets in some grasses are unisexual, and such plants may be monoecious or dioecious. Some plants produce neuter spikelets, often paired with the perfect ones. The positioning of sterile florets and sterile spikelets is of taxonomic significance.

We learn that maturing grass infructescences can be known by the manner in which the inflorescence or spikelets articulate (break naturally into segments). This pairs with archaeological studies that explain selection for non-articulating forms (which did not automatically shed their seed) was a significant development in the domestication of grains.
In order to give you a sample of the way Chase informs us, I’m including several of her illustrations below, annotating each with the notes she provides in her descriptions. Her rationale is perfect; Chase walks us through significant modifications seen in grass inflorescences, explaining and illustrating those very different structures. Take care in reading; the word “spike” refers to an unbranched, tight arrangement of “spikelets”, while the spikelet is the basic flowering unit. Also note that many of her examples are not part of our flora, but they are instructive nonetheless. If only the same extensive treatment existed for all plant families!



In the figures below, Chase illustrates some of the variation found in the lemma, in these cases the lemma is lobed (divided, almost lacerate), each lobe with a nerve (vein) extended into an awn.


In the figure below, Chase describes the inflorescence of Eleusine indica (our non-native Indian Goosegrass) as producing spikes, each crowded with spikelets that overlap in two rows on one side of the rachis, the inflorescence including “two to several spikes borne together, digitate, or nearly so.” Taylor and Flora of North America describe the same plant as bearing panicles. I haven’t encountered a text that offers standards for describing the many grass inflorescence types.



Chase explains this Chloris hybrid as having spikelets with a fertile and sterile floret. Note the sterile floret is stalked. The inflorescence she describes as digitate spikes.


The spikelet of Bouteloua curtipendula (Sideoats Gramma) bears a fertile floret and 1-2 sterile florets. The lemma of the fertile floret and the first sterile floret is awned. Chase describes the inflorescence as a “raceme” because the spikes are spaced along a single rachis. By this standard, the Eleusine, above, could not qualify as a raceme.
The two illustrations below show “fascicles”, tightly branched configurations of spikelets that generate a completely different compounded structure. Note in subsequent figures that Sandspurs (Sandburs, Taylor insists) are a further elaboration of fascicles, resulting from complex stems that surround one or more florets, the stem tips being thorny.




Useful Terminology for Studying Grasses
- Agrostology – the study of grasses, by Agrostologists
- Grassland, Steppe, Llanos, Prairie – biomes dominated by grasses
- Forage – grasses and forbs grown for feedstock
- Runner – equivalent to stolon, a horizontal stem running and rooting along the ground surface
- Stolon – a stem the plays out along the ground, usually anchoring at nodes
- Rhizome – an underground stolon (a running stem), often able to establish a great network of rooted stems
- Fibrous – describing the numerous small tough roots produced by grasses and other monocots
- Sod – a tight mat of grasses, carpeting the ground surface
- Pith – non-vascular tissue that fills the core of a stem; grasses often have a hollow pith, with tissue only at nodes
- Culm – the upright (usually flowering) stem of a grass, from a reedy stalk of Sea Oats to a cane of Bamboo
- Sheath – the basal part of a monocot leaf attaching at a node and enveloping the stem before breaking out to create a blade; grass sheaths are noted as typically split (open) from based to blade
- Blade – the flattened or folded strap of leaf that plays off the sheath
- Ligule – a slight flap or fringe of tissue that marks the juncture of sheath and blade; though usually modest in appearance, the ligule is often an important feature in identification
- Collar – a term describing basal bulging and folding in some leaf blades
- Split – the gap in a grass sheath opposite the axillary bud
- Prophyllum – a rudimentary sheathing leaf associated with emergence of stolons and rhizomes
- Conduplicate vs Involute – contrasting leaf blades that fold along a mid-vein with those that tend to roll in along the parallel margins. Plicate leaves would appear corrugated, folding along several main veins.
- Intravaginal & Extravaginal Branching – contrasts side branches that parallel the main culm, spreading more horizontally above the point the blade peels off from the sheath with branches that pierce the sheath near the node. Rhizomes and stolons normally show extravaginal branching
- Articulate, Disarticulating – refers to the predictable points at which a grass inflorescence will form a suture and break apart at maturity. Important in assuring seed dispersal
- Spike – refers to a straight rachis tightly lined with flowering units (spikelets). Unfortunate that spikelet is so well-defined, while spike is elusive.
- Raceme – refers to a common grass inflorescence type in which the spikes or spikelets are distanced from each other – very unevenly applied, as seems true for most descriptive terms for Grass inflorescences
- Spikelet – the basic flowering unit of grasses (and sedges), usually with two basal bracts called glumes subtending one or more florets
- Rachilla – the slender stem/axis of a spikelet
- Glume – one of two basal bracts common to a spikelet
- Lemma – the most evident and highly variable of two bracts surrounding a flower
- Palea – the inner, lesser, 2-veined bract encasing a grass flower, sometimes adherent to the ovary
- Nerve – what Agrostologists call veins when describing bracts. The nerves of lemmas often extend to become significant awns
- Floret – the term for the grass flower unit, which includes the encasing lemma and palea
- Lodicules– small, seemingly vestigial grass flower tepals with a crucial role in expanding/opening the palea and lemma for dispersal of pollen and exposure of the stigmas
- Awn – an extension of a nerve (typically from a lemma) that makes a serious presence as a pointed, sometimes heavily textured projection
- Rame – an inflorescence branch that may bear both sessile and stalked spikelets, as in the Andropogon and relatives
- Ramet – an ecological term applicable to grasses that spread through stolons and rhizomes, generating functionally separate but connected plantlets, used in scoring abundance
- Distychous – another way to say 2-ranked, describing placement of leaves (phyllotaxy) in grasses, which is always in two lines, alternating along the stem
- Caryopsis – the technical term for a Grass grain, which is a 1-seeded, superior fruit
- Kernel – used for grains important in agriculture, such as maize (called corn in the US) and wheat (called corn in Europe)
- Germ – the developed embry of a grass seed (fruit)
- Endosperm – the bulk storage carbohydrate of many grass seed, a collateral polyploid product of Angiosperm fertilization
- Aleurone – an outer cell layer of the endosperm that generates amylases necessary to break down starch to energize germination
- Pericarp – hardened tissue forming the outer layer of the caryopsis, derived from the fruit wall and thus genetically identical to the parent plants. The pericarp is normally sealed tightly to the seed. In corn, the pericarp is transparent, which is the reason you see expressed color from different genotypes of kernels on a cob of Indian Corn. The pericarp is also the hard, plastic layer that is removed through soaking with lye (nixtamalization), and it’s the part that sticks in your teeth when eating popcorn.
- Mazorca – the Spanish word for an ear, or cob of corn
CYPERACEAE
The Sedges are masters of “reduction”. As with their distant cousins, the Grasses, Sedges have developed basic flowering units called spikelets, but theirs are distinct, as are the “synflorescences” in which flowering spikelets are displayed.
Unsurprisingly, Sedges, noted for their affinity with wet soils, have a strong presence in Florida. With around 140 native species in 13 genera (Bulbostylus, Carex, Cladium, Cyperus, Dulichium Eleocharis, Fimbristyis, Fuirena, Rhynchospora, Schoenoplectiella, Schoenoplectis, Scirpus, and Scleria), we discover there are four genera of substantial numbers: Carex with nearly 80 species, Cyperus with nearly 40, Eleocharis with nearly 30, and Rhynchospora with about 55. The flora also includes another 40 non-native species, most of which are Cyperus.
Most authors tend to describe Sedges in comparison to their more familiar cousings, the Grasses, which proves useful since making determinations in these plant groups can be difficult and they are often confused with one another – making for seemingly unsurmountable challenges. There are, however, many clues as to when a plant is a Sedge as opposed to a Grass (generally reliable, but not absolute).
If you locate a monocot with 3-ranked leaves, especially if the foliage rides along a 3-sided (triangular in x-section) culm (aerial stem), that’s almost certainly a Sedge, especially when it shows a sharp edge (“sedges have edges…”). The only time I’ve been confused has been when spotting tufts of Zygadenus glabberrimus foliage, which also show this “triquetous” (triangular) phyllotaxy – but a second glance at the showy flowers and round scapes of Zygadenus quickly dispatches any such thoughts. Examining the leaf bases of Sedges, you’ll find the sheathes completely surround the stem; in Grasses, the sheath is commonly split to the node. Cutting a Sedge stem, you’ll find the pith filled with cells, which isn’t the case for all Grasses (most of which have hollow internodes).
Sedges differ from most Grasses in bearing leafy bracts at the bases of their flowering structures (“synflorescences”), which Vegetti (2003) describes as based on one of three sorts: the Panicolodium (panicle-like), the Antholodium, and the Truncated Antholodium (which lacks a central, i.e. terminal, fertile spikelet). See his illustration below:

Knowing something about sedges, this doesn’t quite jive with expectations. What about Eleocharis, with just a single spikelet? And those plants with with tight heads of spikelets? Vegetti explains the rest is a simple matter of contraction and reduction. Shorten all internodes in his anthelodium and you have a head. Eliminate all but the terminal spikelet, and you have Eleocharis – the Spike Rushes.
But that’s just a taste of reduction that’s been achieved by Sedges. Check out the extraordinary diagram created and published by Ilkka Kukkonen (1984). I marvel at his patience and tenacity in working out this diagram.

As was mentioned earlier, Sedges bear their flowers in bract-built spikelets, the organization of which might be distichous (2-ranked and basically planar, similar to those of grasses) or tightly spiraling. As in Grasses, Cyperaceous spikelets themselves are hyper-reduced inflorescences, and variable in composition.
Most scales in a spikelet will support fertile flowers flowers, which will be perfect (both pistil and anthers) or imperfect (either male or female); flowers of Carex (the largest genus) and Scleria are all imperfect, male and female flowers often present in the same spikelet. The flowers are practically naked, some bearing slender filaments interpreted as relictual perianth segments. Pistillate flowers bear a sole superior, single-seeded ovary that matures as an achene (also called a nutlet).

To key out the identification of a Sedge, be prepared with flowering and fruiting material and a handlens or dissection microscope. You’ll be jumping into the fire, with determinations dependent on whether the flower has a perigynium, the number and character of any filamentous perianth segments that are present, the nature of any projection at the summit of the achene, etc. Most of the nearly 50 couplets to genera of Cyperaceae in Flora of North America involve close inspection of spikelets and flowers.
Terminology Useful for Cyperaceae
- Caespitose – in tufts, tightly clustered, kept close to the soil, as in tight clumps of short leafy stems we see in many sedges.
- Perigynium – a scale-like structure that encloses flowers of Carex and a few other genera, separate from the spikelet scale
- Perianth – tepals or segments present in flowers of some genera, usually filamentous. Those of Scirpus conspicuously curled, wooly; numerous in Eleophorum; absent in Cyperus, Cladium, and Fimbristylis
- Bristle – term for dried perianth segments when they remain attached to the mature achene
- Tubercle – a prominence on the achene (fruit) formed by the persistent base of the style. You’ll need to appreciate the shape and extent of a tubercle for identification of certain species.
Eudicots
The Angiosperm Phylogeny Group has developed patterns of exclusion and inclusiveness – using terms like Core, Super, and True (Eu). Thus there are Eudicots (true Dicots, which fold in families like the Ranunculaceae, Papaveraceae) and Core Eudicots (really, truly Dicots, the groups that settled on a set number of petals, i.e. a lot of our Wildflowers). Among our native plants we find representation in those Eudicots not on the varsity team (not Core Eudicots) – the Boxwoods, Proteas, Sycamores, and Ranunculids. Here’s the breakdown:
- Buxales: Buxaceae (Native 1/1 Pachysandra)
- Proteales: Nelumbonaceae (Native 1/1 Lotus; Exotic 1/1 Lotus), Platanaceae (Native 1/1 Platanus)
- Ranunculales: Berberidaceae (Native 1/1 Podophyllum; Exotic 2/2 Nandina, Mahonia), Menispermaceae (Native 4/4 Calycocarpum, Cissampelos, Cocculus, Menispermum), Papaveraceae (Native 3/5 Argemone, Corydalis, Sanguinaria; Exotic 2/5 Fumaria, Papaver), Ranunculaceae (Native 8/23 Actaea, Anemone, Aquilegia, Clematis, Delphinium, Enemion, Ranunculus, Thalictrum, Xanthohiza; Exotic 3/9 Clematis, Delphinium, Ranunculus)

Many of our wildflowers are among the plants considered True Dicots (Eudicots). I’ve indicated Orders, but include a reminder here that current wisdom suggests three groupings, the basal Eudicots we just discussed (Poppies, Lotus, Clematis), and two major clades, the Rosids and Asterids. Families discussed in greatest detail include Fabaceae, Euphorbiaceae, and Onagraceae as representing the Rosids, with Asterids exemplified by Lamiaceae, Apocynaceae, Convolvulaceae, and Asteraceae.
It’s not so simple, of course. We find the botanists hedging bets, in that both Rosids and Asterids are qualified; there’s the SuperRosids and Rosids, paralleled by SuperAsterids and Asterids. In the case of SuperRosids, systematists tuck plants we know as Sedums, Saxifrage, Itea, Sweetgum and Witch Hazel close to the Rosids, and with SuperAsterids, they position the Caryophylls (from Phytolacca to Polygonum to Cacti) and the Mistletoes somewhere alongside the Asterids. This tethers the Caryophylls, long recognized as a defined clade (based on many characters), to the Asterids, suggesting this branch is not as distinct from other Dicots as we we were taught in the past.
In the next-to-last (penultimate) section, we begin by examining the Rosids, which are gathered into three loose associations: the everyday Core Fabids (also called Fabids and N-Fixing Fabids), the COM (an acronym for Celastrales, Oxalidales, and Malpighiales) clade, and the Malvids. Let’s review the families included.
SUPERROSIDS & VITALES
- Saxifragales: Altingiaceae (Native 1/1 Liquidambar), Grossulariaceae (Native 1/1 Ribes), Haloragaceae (Native 2/5 Myriophyllum, Proserpinaca; Exotic 1/2 Myriophyllum) Hamamelidaceae (Native 2/3 Fothergilla, Hamamelis; Exotic 1/1 Loropetalum) Iteaceae (Native 1/1 Itea), Penthoraceae (Native 1/1 Penthorum)
- Vitales: Vitaceae (Native 5/11 Ampelopsis, Cissus, Nekemias, Parthenocissus, Vitis; Exotic 2/3 Causonis, Cissus)



Though predominately woody, we can’t completely ignore the “Superrosids” (also Vitales) which include a few herbaceous representatives, the aquatics Myriophyllum and Proserpinaca. More importantly, though, anyone prospecting for native plants in Florida will have faced the veil of Vitis rotundifolia tangled with cordons of prickly Smilax that enmesh practically any moist or wet shrubby edge. It’s too easy to dismiss Florida’s biomass of tough vines when discussing habitats, but they are the reason the word “impenetrable” might be needed in natural history essays.
The EUROSIDS
The Eurosids (True, or Core Rosids) break into three main groups, the Fabids, the COM-Clade, and the Malvids. We talk about one family in each group, the Fabaceae in the Fabids, the Euphorbiaceae in the COM-Clade, and the Onagraceae among the Malvids.
FABIDS & N-FIXING+ FABIDS
- Zygophyllales: Krameriaceae Native 1/1 Krameria), Zygophyllaceae (Native 2/2 Guaiacum, Kallstroemia; Exotic 3/4 Guaiacum, Kallstroemia, Tribulus)
- Cucurbitales: Cucurbitaceae (Native 4/5 Cyaponia, Cucurbita, Melothria, Sicyos; Exotic 9/15 Citrullus, Coccinia, Cucumis, Cucurbita, Lagenaria, Luffa, Momordica, Sicyos, Trichosanthes)
- Rosales: Rosaceae (Native 11/32 Agrimonia, Amelanchier, Aronia, Crataegus, Fragaria, Malus, Physocarpus, Potentilla, Prunus, Rosa, Rubus; Exotic 9/17 Aphanes, Eriobotrya, Photinia, Potentilla, Prunus, Pyracantha, Pyrus, Rosa, Rubus), Rhamnaceae, (Native 9/12 Berchemia, Ceanothus, Colubrina, Frangula, Gouania, Krugiodendron, Pseudoziziphus, Reynosia, Sageretia; Exotic 2/3 Colubrina, Ziziphus), Ulmaceae (Native 2/5 Planera, Ulmus; Exotic 1/1 Ulmus), Urticaceae (Native 5/11 Boehmeria, Laportea, Parietaria, Pilea, Urtica; Exotic 8/8 Boehmeria, Cecropia, Laportea, Parietaria, Pilea, Pouzolzia, Rousselia, Urtica), Cannabaceae (Native 2/6 Celtis, Trema; Exotic 2/3 Cannabis, Trema), Moraceae (Native 2/3 Ficus, Morus; Exotic 8/18 Artocarpus, Brosimum, Broussonetia, Dorstenia, Fatoua, Ficus, Maclura, Morus)
- Fagales: Betulaceae (Native 4/4 Alnus, Betula, Carpinus, Ostrya, Fagaceae (Native 3/29 Castanea, Fagus, Quercus plus hybrids; Exotic 1/2 Castanea), Juglandaceae (Native 2/8 Carya, Juglans, Myricaceae (Native 1/3 Morella (Myrica))
- Fabales: Fabaceae, Polygalaceae (Native 2/23 Asemeia, Polygala (Senega), Surianaceae (Native 1/1 Suriana)
Everyday Fabids include Orders that populate our gardens, forests, and waysides – the Rosales (Roses, Elms, and Figs), Fabales (Legumes and Polygalas), Cucurbitales (Cucurbits and Begonias), and the wind-pollinated Fagales (Oaks, Walnuts, Casuarinas, and Bayberries). This grouping is, to me, quite the mixed bag.
The Rosales cluster Rosaceae, a family of plants we imagine as producing textbook flowers (with 5 sepals, 5 petals, numerous stamens, and separate carpels) and the Rhamnaceae (with their sweet clawed petals) are not the full story.

Most of our native Rosaceae are woody, while Agrimonia, one of our few native herbaceous Rose family natives, differs in various regards. With insignificant sepals and petals, flowers are a bit more similar to the curious assembly of related families with highly reduced floral structure, the Ulmaceae, Cannabaceae, Moraceae, and Urticaceae. But for the nettles, these families comprise woody plants. Paralleling that is a grove of trees (Oaks, Walnuts, Hickories, Beeches) also showing highly-reduced flowers, an assembly we used to call the Amentiferae, referencing their male flowers borne in catkins (tassels). Once again, though native, these are woody plants, mostly forest trees.
Within the Rosids, Fabales stands out as the Order most prominent among Florida’s Wildflowers, well populated with native herbaceous Wildflowers (many Fabaceae and all of the native Polygalaceae).
FABALES
FABACEAE

When I say let’s get into the Legumes, I wonder how much meaning the word “legume” has with a wildflower audience. I think of “legume” as a common word people associate with peas and beans, at least with any plants that produce a peas-in-a-pod fruit. So I’m unclear as to how much mileage I can expect with a “cold ask” to explore the Legume family (the Pea Family, the beans, the Fabaceae) before we encounter the unfamiliar.
It’s unlikely we need to go through superlatives, vaunting the protein content of legumes in diets, because these plants are known champions. Certainly soybeans, an exotic novelty when I was a child, have broken marketing and market barriers, becoming a significant crop for American farmers and entering our diets in a range of products, not simply the salt-laden sauces that introduced me to American-style Chinese cuisines. Having grown up in Lower Alabama, I was well-acquainted with Peanuts, Arachis hypogaea, which we inherited from peoples of the Andes and South America along with other incredible underground food sources, like Potato (Solanum tuberosa – Solanaceae), Oca (Oxalis tuberosa – Oxalidaceae), Yacon (Smallanthus sonchifolius – Asteraceae), and Ahipa (Pachyrhizus ahipa – Fabaceae).








We are also familar with legumes in the landscape. Any gardeners in warm climates like Florida’s will know the exotic and indestructible Peacock Flower, the Pride of Barbados, Caesalpinia pulcherrima, or the Powderpuff, Calliandra. Between Peanuts and Peacock Flowers, two commonly cultivated plants, we reconcile our grasp on the family – suddenly it’s “by their fruit ye shall know them.” I believe most plant people accept legumes as plants that, despite differences in agreement as to how the flower might be shaped, are all determined to make a single pod with something like a pea or bean seed inside.

We are led to the observation there are a few basic flower types that characterize Legumes one encounters in Florida. We find: 1. the “pea” type flower, with a large petal (the ‘standard’ or ‘banner’), two that form a ‘keel’ concealing the stamens and single ovary, and two ‘wings’ that hang out on the sides (i.e Wisteria, Clover, Centrosema); 2. the “medallion” type, with a flat flower and short stamens, like Cassia and Chamaecyperus. 3. the “petal & anther” forms, with showy petals and showy anthers (Peacock Flower); and 4. the “powderpuff “type, that have showy anthers and reduced petals (Mimosa, Acacia).


But once again, we are deceived by our “lying eyes.” The Fabaceous world of my botanical innocence was a land of fable, at least when I investigate current studies and dogma. You are welcome to follow along by visiting the Legume Data Portal, a collaborative project across the realm of people who study systematics in the Fabaceae (the Legume Phylogeny Working Group, LPWG), where summaries and indices covering a range of topics, most specifically in the realms of taxonomy and systematics, are posted.
Examining the Portal, I find my primordial assumptions, my pre-cladistics-era outline of three distinct and easily distinguishable groups of Legumes (the Fabs, the Mimosas, and Caesalpinoids), like the Federal Government, has been dismantled, relegated as artifact. I wish I understood the character states and cladistic wizardry behind the current schema, but I can only serve as messenger. Here’s the scoop:
Worldwide, about 800 Legume genera and over 23,000 species are recognized. Current wisdom allocates those plants to six subfamilies: Cercidoideae, Detarioideae, Duparquetioideae, Dialioideae, Caesalpinioideae, and Papilionoideae. You can ignore the three D-groups, the unfamiliar Detarioids, Duparquetioids, and the Dialioids, which are smaller, tropical segregates. Duparquetioideae is monotypic; only a single species is recognized. Sidelining those spares has meant little to us in Florida. Those plants are a long way from home, in a galaxy far, far away.
Better-known plants have been reshuffled however. Subsuming the Cassias (Medallion-type) and the Acacias (Mimosoid) with the Caesalpinoids has required some mental adjustment, I can deal with that. But learning that Cercis, our Redbud, with its fairly-normal seeming “pea” flower is a headliner for a separate subfamily (Cercidoideae, which also includes Bauhinia) could bear some explanation, as does learning Pisum is no longer considered an accepted genus, having been subsumed to Lathyrus. What would Mendel say?
In Florida, the ISB Plant Atlas lists nearly 100 genera boasting 340 taxa. Of those plants, records include just a single plant in the three D-groups, the non-native Tamarindus from the Old World tropics. That leaves us with the Cercidoids, the Caesalpinioids, and the Papilionoids to consider. Here’s a summary:
The flora lists twenty-seven genera of Legumes in the Caesalpinoids, fourteen of which are not native. Exotics include the woody Acacia, Albizia, Calliandra, Peltophorum, and Parkinsonia. Native Caesalpinoids include herbaceous Chamaecrista and Mimosa species, and several Senna.

The flora records three genera of Cercidoids in Florida: Bauhinia, Phanera, and Cercis; of these only the Redbud, Cercis canadensis is considered native.

That leaves the bulk of our Legumes (65 genera) as Papilionoids (pea-flowered). Twenty-six of those genera are totally non-native, most particularly Medicago and Melilotus, as well as the bulk of Trifolium, Crotolaria, Indigofera, and Wisteria species.



In summary, of 97 genera of Legumes in the flora, only twenty genera are purely represented by native plants. Forty-four genera are totally exotic, leaving thirty-two genera of mixed status, with some native and some non-native representatives. These numbers speak to the significance of non-native Legumes in the state. It’s also clear the great bulk of Legumes that would be considered native Wildflowers are Papilionoids – herbaceous to suffrutaceous plants with pea-flowers, such as Apios, Astragalus, Baptisia, Centrosema, Clitoria, Dalea, Desmodiuim, Galactia, Lespedeza, Lupinus, Strophostyles, Stylosanthes, and Tephrosia.
Terms Useful in Studying Legumes
- Legume – used in two senses: 1. a particular kind of pod that dehisces along a single suture, with seed along a linear
- Loment – a particular kind of pod in which the fruit wall constricts, even segments, between individual seed – as in Desmodium and Sophora.
- Stipule – the foliar tissue, usually paired, at the juncture of leaf petiole and stem – a standard character for many plant groups, but particularly significant among Legumes, with a leafy presence in peas varying to hollow spines on some Acacias.
- Hypanthium – as used for Legumes, an almost inconspicuous aspect of Fabaceous flowers, with a modest floral cup yielding to a cup-shaped calyx – not to be confused with hypanthium as used to describe Oenothera and other plants with clearly inferior ovaries
- Standard (Banner) – the large petal in Faboids opposing the positioning of stamens and pistil wrapped in the paired keel petals. In some plants, the flower is “resupinate” (repositioned by 180 degrees) and Standard is therefore presented lowermost rather than uppermost
- Wing – each of two lateral petals between the Standard and Keel
- Keel – a boat-shaped structure formed of two petals and surrounding the stamens and pistil, directly engaged in pollination mechanisms
- Monadelphous – stamen filaments fused to form a single structure, tubular or not
- Diadelphous – stamens with filaments fused (connate) to form two groups or fascicles, in Legumes often a group of many and a single unfused stamen
- Gynophore – a stalk that lifts/extends the ovary from it’s attachment to the receptacle
- Hilum – a visible abscission zone where a seed was attached to the funiculus (basically a belly-button) that was the developmental lifeline to the placenta. Characteristics of the hilum are readily noticeable in beans and peas.
- Compound leaves – Legumes are noted for various levels of compound leaf blades, from ternate, to evenly- and oddly-pinnate or bipinnate blades.
- Pulvinus – a swelling present at the base of petioles (even petiolules) of Legumes, actively engaged in leaf movements
- Thigmonastic – movement of leaves and other plant parts stimulated by touch
- Root Nodules – Specialized masses of root tissue that provide low-oxygen housing for Nitrogen-fixing bacteria – typical of many Legumes, granting those plants superpowers as keys to localized soil enrichment and generators of seed and plant mass known for their protein-rich potential.

COM-CLADE
Three Orders are thought to form a natural branch, a clade within the Rosids, simply called the COM clade because it comprises the Celastrales (think Parnassia), the Oxalidales, (i.e. Oxalis) and the sizable Malpighiales (Hypericum, Linum, Passiflora, Violets, and the Euphorbs). I introduced Hypericum and Viola in earlier, separate treatments, so for this discussion let’s delve into the Euphorbs, a family showing extreme levels of floral modification and reduction
- Celastrales: Celastraceae incl. Parnassiaceae (Native 7/10 Crossopetalum, Euonymus, Gyminda, Hippocratea, Parnassia, Schaefferia, Tricerma; Exotic 2/2 Celastrus, Euonymus)
- Oxalidales: Oxalidaceae (Native 1/3 Oxalis; Exotic 1/5 Oxalis)
- Malpighiales: Chrysobalanaceae: (Native 2/2 Chrysobalanus, Geobalanus), Clusiaceae (Native 2/35 Hypericum (34 taxa), Clusia; Exotic 1/2 Calophyllum), Euphorbiaceae (Native 12/68 Acalypha, Argythamnia, Astraea, Caperonia, Cnidoscolus, Croton, Ditrysinia, Euphorbia, Gymnanthes, Stillingia, Hippomane, Tragia; Exotic: 17/35 Acalypha, Aleurites, Caperonia, Cnidoscolus, Codiaeum, Croton, Dalechampia, Euphorbia, Hura, Jatropha, Mallotus, Manihot, Microstachys, Ricinus, Sapium, Vernicia), Linaceae (Native 1/9 Linum; Exotic 1/2 Linum), Malpighiaceae (Native 1/1 Byrsonima; Exotic 5/6 Galphimia, Heteropterys, Hiptage, Malpighia, Stigmaphyllon), Passifloraceae incl. Turneraceae (Native 2/7 Passiflora, Piriqueta; Exotic 2/8 Passiflora, Turnera), Phyllanthaceae (Native 2/4 Hetersavia, Phyllanthus; Exotic 4/11 Bischofia, Breynia, Flueggea, Phyllanthus), Putranjivaceae (Native 1/2 Drypetes), Rhizophoraceae (Native 1/1 Rhizophora; Exotic 1/1 Bruguiera), Salicaceae (Native 3/8 Populus, Salix, Xylosma; Exotic 6/6 Casearia, Flacourtia, Oncoba, Populus, Salix, Xylosma), Violaceae (Native 2/10 Hybanthus (Cubelium), Viola; Exotic 2/4 Pombalia, Viola)
MALPIGHIALES
EUPHORBIACEAE

Encountering a plant with white latex, or milky sap, I run through a mental checklist. Does it have opposite leaves (i.e. Apocynaceae)? Is there an encircling scar at the node, where fused stipules had been attached (Moraceae)? Is it a Composite, like Lactuca or something similar? Are any flowers present that would point to Campanulaceae, or even the Convolvulaceae? Or, most tellingly, if the leaves are alternate, do I find a fruit that’s evidently 3-lobed? If so, the plant in hand is likely a Euphorb – and I’m reminded to keep hands away from my eyes. The latex is likely caustic.
Indeed, the Euphorbs are full of bad news when it comes to toxins and chemistry, something I learned from an encounter with Hura crepitans in Panama decades ago. The family is well-known for poisons; you’ll discover the established names for plants reflect an association with toxic compounds, following an ancient concept that the cure is in the dose. Euphorbia was named for Euphorbus, an early Greek physician. Jatropha is derived from Greek terms for physic and food, given a Mediterranean species was used historically as a purgative. Cnidoscolus combines Greek words for nettles and stinging hairs. Tragus (called Noseburns) commemorates the early German herbalist Hieronymus Bock, whose surname Latinizes as tragus (goat). Linnaeus derived Hura from a native American word for the poisonous sap (something to which I can attest). Manihot comes from a native Brasilian term for yuca (the original meaning implying a “wood spirit”), which is the root a plant, some forms of which must be purged of toxins to generate tapioca. Acalypha, though not an irritant, still has a name referencing the Stinging Nettles (Urtica) due to similarity in the foliage. It’s a potent group.
The family, broadly defined, is characterized by unisexual flowers. In some genera, Jatropha and Cnidoscolus, flowers form showy perianths (both sepals and petals in Jatropha, sepals-only in Cnidoscolus) – almost “normal.” Several other genera, particularly Croton, Stillingia, and Tragia have less-conspicuous male and female flowers, with reduced or absent perianths. Pistillate flowers in these plants are basically naked, perched, nearly camouflaged along the inflorescence axis (typically basal).
In Euphorbia and related genera, the perianths are not developed, and flowers reduced to the point that what is technically a unisexual flower is reduced to a single reproductive element, such as one anther or one naked ovary. Through reducing the flower to its core functions, we discover the plants have evolved great flexibility in re-amalgamating sexual components to create a range of pseudo-flowers (pseudanthia) that can take advantage of various pollinating vectors.
With the Sedges and Grasses, we encountered highly reduced flowers and condensed inflorescences as spikelets and compounded inflorescences. Among the Dicots, we also see natural families characterized by reduction and aggregation of flowers. In Composites, which we’ll examine later, the head or capitulum is a pseudanthium of reasonably straight-forward character. The Euphorbs, among Dicots, show extremes of floral reduction and aggregated inflorescences in a glorious buffet of elaborations. They are the masters of floral reduction (surpassing the Moraceae), which means interpreting flowers, even determining where one flower ends and the next begins, can be challenging. Thankfully, a consortium that was active until 2012, the Euphorbia Planetary Biodiversity Inventory project, has left a legacy website that gives access to illustrations and information concerning Euphorbia, the most significant genus in the family (as well as in our flora), and one of the largest genera of flowering plants, with over 2,000 species
The reduced Euphorbia flowers are organized in a variable structure, the cyathium (see below), which consists of an involucral cup that at its simplest might integrate one or more glands along with an ovary and a stamen (each being a separate reduced flower). In Poinsettia (Euphorbia pulcherrima) and others, we find those cyathia compounded in a flowering structure that includes colorful bracts, now a complex synflorescence. The related Pedilanthus (currently folded into Euphorbia) includes cyathophylls associated with hummingbird pollination.


At a very different end of the spectrum, the Acalypha contingent has explored wind pollination, with fascicled, multiplied stamens and branched stigmas. Glands in some Dalechampia flowers (in the Tropics) generate essential oils that attract Euglossine bees as vectors.
Checking data on the The Florida flora in ISB, you’ll see 24 Euphorbiaceous genera, 12 of which are introduced, including trees such as Vernicia (Aleurites), Hura, and Sapium (Triadica) the various shrubby Jatropha, the multicolor-leaved shrub (Codiaeum), the exuberant wayside Ricinus, and vining Dalechampia.



Most Croton and Euphorbia species listed are native, but each genus includes several introduced taxa. Both species of Stillingia and all of the nettlesome Tragia are considered native.
Among native Euphorbs, practically all of which are herbs or small shrubs, Cnidoscolus and Caperonia stand out with their conspicuous white flowers (which in Cnidoscolus are considered sepals, sometimes called a “calycine perianth”), adapted to moth pollination.

The bulk of native Euphorbaceae are in the genus Euphorbia. Among those, some are easier to spot than others. The plant I call Wild Poinsettia, Euphorbia cyathophora, stands out with red-splotched leaf bases surrounding a terminal cluster of cyathia. It’s reported as native, though I seem to encounter specimens near residences and not in wilder settings.

A pair of distinctive plants encountered frequently in the Panhandle are the two Stillingias. Stillingia sylvatica (which for a reason I’ve never understood is called Queen’s Delight) shows up in sandy soils of pine flatwoods, while the wetland S. aquatica, with its characteristic form (a top-branched, single-stemmed subshrub) and sickly yellow-green foliage seems ever-present in shallow standing waters.
Terminology Useful in Studying Euphorbs
- Laticifer – specialized cells producing latex, used when the cells tend to form tubular canals
- AGI – Alkaloidal Glycosidase Inhibitors, bioactive compounds collected by Uraniine moths
- Malpighiaceous trichomes (hairs) – an unusually-structured trichome described from plants in the Malpighiaceae, but now known in various plant families – characterized as a flattened, sometimes branching trichome that is held tight to the epidermis by attachment at or near the center of the cell.
- Cyathophyll – The small, colorful bracts attached to the base or rim of cyathia, resembling the petals of a normal flower. These are distinct from the colored leaves seen below inflorescences in Poinsettia and Wild Poinsettia
- Anthocladia – leafy, repeating chains of inflorescences characteristic of various plant groups
- Petaloid Calyx – the form of Cnidoscolus flowers, in which the white “perianth” is really a “calyx” – reminiscent of the unrelated Nyctaginaceae – same as Calycine perianth.
- Pseudanthia – in Euphorbs, the combination of features that causes a cluster of highly-reduced flowers to to appear as a single flower – sometimes equivalent to the term Cyathium, but seemingly applied more extensively, as in the pseudanthium of Dalechampia (which can be Euglossine pollinated)
- Pleiochasium – an inflorescence that generates a cluster of flowering structures that mature simultaneously, similar to cauliflower
- Synflorescence – used variably, appllicable when several highly reduced structures are closely gathered
- Allomorphic Pistillate Flowers – In Acalypha, distinct pedicellate flowers with basal styles borne among more normal pistillate flowers, species-specific
- Calycine perianth – a showy perianth composed of sepals rather than petals
- Involucral cup – the slightly concave stem tip that gathers reduced flowers into a cyathium
- Cyathium – the swollen stem tip that forms an involucral structure holding stamens and a pistil, each of which, morphologically, is a single highly reduced flower. The Cyathium usually includes one or more glandular structures, and may be advertised by petal-like bracts called cyathophylls.
- Apetalous – a flower structure lacking petals, which is most of the family
- Dissected style – highly branched styles in the wind-pollinated Acalypha-related (Acalyphoid) Euphorbs
- Androecium – referencing stamens when there is some common particular structural character to discuss
- Fascicled Stamens – Stamens united in groups (versus “free”) that branch, proliferating the number of anthers, feature of wind-pollinated Euphorbs, Ricinus and other Acalypha relatives
- Anther position (Erect vs. Inflexed, Inclinate, Basifixed, Sessile) -characters observed and reported in anther development as buds mature
- Inaperturate, Crotonoid – Pollen showing no apertures, particular to Croton and relatives
- Carpidiophore – a persistent, triangular woody base to the fruit of Stillingia
- Schizocarp – fruit type that dries and dehisces (splits open) explosively
- Pachychalazy (Pachychalazal) – a characteristic noted in various seed plant families (Euphorbiaceae, Rutaceae, Anacardiaceae) in which the chalazal cells (cells at the distil end of the egg sac) develop into thickened tissue, sometimes apparent as a component of the seed coat.
- Aril – fleshy tissue produced on the surface of seed, a food source typically promoting in dispersal
- Caruncle – an aril produced around the hilum/micropylar region of the seed coat in Euphorbs, functioning as an elaisome that attracts ants
- Sarcotesta – the fleshy seed coat of many tropical Euphorbs
- Exarillate – lacking an aril, in a plant group otherwise expected to produce aril
The MALVIDS (Brassicales, Malvales, & Myrtales)
The Malvids encompass a host of important wildflower groups, most especially the Malvales (Cistaceae (Crocanthemum and Lechea) and Malvaceae, with Hibiscus and relatives) and the Myrtales (the Lythraceae, Melastomes, and Onagraceae).
Checking out Cistaceae (Malvales) in the Florida Plant Atlas, we see only two genera represented, Crocanthemum (with six species) and Lechea (with nine species), all of which are native to the state. The two genera key out based on width of leaves and breadth of flowers. Lechea leaves are threadlike, less than a millimeter wide, and the flowers are similarly miniature, less than 2 mm across. No wonder the Lecheas are called Pinweeds. The endemic Lechea lakelae is reported only from Marco Island, in Collier County.



- Brassicales: Bataceae (Native 1/1 Batis), Brassicaceae (Native 10/24 Abdra, Boechera, Cakile, Cardamine, Descurainia, Lepidium, Nasturtium, Planodes, Rorippa, Tomostima, Warea; Exotic 15/21 Brassica, Capsella, Cardamine, Cleoserrata, Conringia, Diplotaxis, Erucastrum Erysimum, Lepidium, Lobularia, Nasturtium Raphanus, Sinapis, Sisymbrium, Thlaspi, Capparaceae (Native 2/2 Cynophalla, Quadrella), Caricaceae (Native 1/1 Carica), Cleomaceae (Native 1/1 Polanisia; Exotic 4/5 Arivela, Gynandropsis, Sieruela, Tarenaya)
- Crossosomatales: Staphyleaceae (Native 1/1 Staphylea)
- Geraniales: Geraniaceae (Native 1/2 Geranium; Exotic 2/3 Erodium, Geranium)
- Malvales: Cistaceae (Native 2/15 Crocanthemum, Lechea) Malvaceae (Native 17/64 Abutilon, Ayenia, Callirhoe, Cienfuegosia, Corchorus, Gossypium, Herissantia, Hibiscus, Kosteletzkya, Malachra, Malvastrum, Modiola, Pavonia, Tilia, Waltheria, Melochia, Sida; Exotic 21/42 Abelmoschus, Abutilon, Anoda, Ayenia, Callirhoe, Firmiana, Grewia, Hibiscus, Malachra, Malva, Malvastrum, Pavonia, Thespesia, Wissadula, Corchorus, Malvaviscus, Melochia Talipariti, Urena,Triumfetta, Sida), Thymelaeaceae (Native 1/1 Dirca)
- Myrtales: Combretaceae (Native 3/3 Conocarpus Lugunaria, Terminalia; Exotic 3/6 Combretum, Lumnitzera, Terminalia), Lythraceae (Native 6/11 Ammannia, Cuphea, Decodon, Didiplis, Lythrum, Rotala; Exotic 3/5 Cuphea, Lagerstroemia, Rotala), Melastomataceae (Native 2/11 Miconia, Rhexia; Exotic 3/3 Acisanthera, Heterotis, Melastoma), Myrtaceae (Native 4/8 Eugenia, Mosiera, Myrcia, Myrcianthes; Exotic 8/16 Corymbia, Eucalyptus, Eugenia, Melaleuca, Pimenta, Psidium, Rhodomyrtus, Syzygium, Onagraceae (Native 2/37 Ludwigia, Oenothera; Exotic 2/9 Ludwigia, Oenothera)
- Picramniales: Picramniaceae (Native 2/2 Alvaradoa, Picramnia)
- Santalales: Olacaceae incl. Ximeniaceae (Native 1/1 Ximenia), Santalaceae incl. Viscaceae (Native 1/2 Phoradendron; Exotic 1/1 Santalum), Schoepfiaceae (Native 1/1 Schoepfia)
- Sapindales: Anacardiaceae (Native 3/8 Metopium, Rhus Toxicodendron; Exotic 4/4 Mangifera, Schinus Sorindeia, Spondias), Burseraceae (Native 1/1 Bursera), Meliaceae (Native 1/1 Swietenia; Exotic 4/5 Azadirachta, Khaya, Melia, Swietenia), Rutaceae (Native 3/8 Amyris, Ptelea, Zanthoxylum; Exotic 5/6 (+ 5 hybrid Citrus) Atalantia, Citrus, Glycosmis, Murraya, Triphasia), Sapindaceae (Native 8/15 Acer, Aesculus, Cardiospermum, Cupania, Dodonaea, Exothea, Hypelate, Sapindus; Exotic 8/11 Blighia, Cardiospermum, Cupaniopsis, Dimocarpus, Harpullia, Koelreuteria, Melicoccus, Sapindus), Simaroubaceae (Native 2/2 Lietnera, Simarouba; Exotic 1/1 Ailanthus)

Examining the list of Florida Malvaceae, we see nearly 80 species represented, slightly more than half of which are non-natives. Among the native Malvaceae, many are restricted to the southern counties, such as Cienfuegosia (reported only from Key West), Avenia, Malachra, and Waltheria. The largest genus, Hibiscus, shows seven native and eight exotic species. Of the natives, three are northern (Hibiscus laevis, H. aculeatus, and H. moscheutos), two are southern (Hibiscus furcellatus and H. poeppigii), and the remaining couple (Hibisus grandiflorus and H. coccineus) range throughout. There are few truly herbaceous native Malvaceae, Callirhoe being the most prominent.

Still within the Malvid clade, we move to another other cluster of families, the Myrtles, plants that flaunt a noteworthy, sculptured hypanthium, that is, plants that show off well-developed inferior fruit, often with very obvious, usually persistent triangular calyx lobes. The woody native representatives of this Order in our flora are predominately Myrtaceae (Eugenia, Mosiera, Myrcia, and Myrcianthes), genera with tropical centers of diversity. Among introduced Myrtaceae, count Psidium (Guava, with its swollen inferior fruit) and the invasive Melaleuca, competing with Pythons to own the Everglades. Our significant wildflowers fall out in three related families, the Melastomataceae, Lythraceae, and Onagraceae.
Native Melastomes, though present throughout the state, are represented by two genera only, Miconia (Miconia bicolor is reported from Miami-Dade County only) and Rhexia, which is represented by ten native species, with a particular distribution pattern. Two species (Rhexia salicifolia and R. parviflora) are reported from the western Panhandle, a cadre of distinct plants are reported from the Panhandle and the northern Peninsula (Rhexia alifanus, R. nuttallii, and R. petiolata), while Rhexia mariana, Rhexia nashii, and R. cubensis are reported from practically every region of the state, with the exception of the southernmost tip (which is the very area that harbors Miconia bicolor.) This link to Rhexia, takes you to a discussion of that genus as I have encountered plants in the Apalachicola region.

I’ve also given some time to understanding heterostyly in local Lythrum populations, but need to pay more attention to these herbs. Reviewing native Lythraceae (and their distribution), it seems they reflect a similar pattern to those described for Rhexia and Hibiscus. The Atlas shows eleven species (in six genera). Two are endemic – our native Cuphea aspera, described from Gulf County, west of Apalachicola by Chapman, and Lythrum nieuwlandii (a plant described a century back by Julius A Nieuwland, and only recently rescued from taxonomic purgatory by Franck and Werner) – known only from Alachua, Citrus, and Hernando counties. The endemic Lythrum flagellare isreported from South Florida only. The near-endemic Lythrum curtissii shows that curious disjunct pattern around the Suwannee embayment (Apalachicola to the west and the Atlantic coast to the East). Willow herb (Decodon) is present throughout, but for the more tropical southern area, while Lythrum lineare has been collected in coastal counties throughout. The aquatic Didiplis is known from Louisiana and Alabama, but is represented in our flora only by collections from Sarasota County. Finally, we have Ammannia, which seems to occur throughout the state.


Based on pure numbers, Myrtalean wildflowers are reflected by more species of Ludwigia and Oenothera than in related families, thus we focus on this group. You’ll note the Onagraceae share many characters with Lythraceae and Melastomataceae, most particularly a pronounced hypanthium topped by a crown of stiffly-distinct calyx lobes, the inferior fruit of Lythrum not so dissimilar from that of Ludwigia, and the 4-petaled Rhexia echoing that standard seen throughout Onagraceae.
MYRTALES
ONAGRACEAE
Worldwide, Onagraceae is not a large group, a family with 22 genera and under 700 species. Taxonomically, we’re looking at a well-studied and documented family, known for PTH (Permanent Translocation Heterozygosity), a chromosomal condition that allows individuals to self-procreate while avoiding the normal deleterious impacts of inbreeding.
Treatments uniformly segregate Ludwigia (the genus in our flora with the largest number of species) in a subfamily distinct from the remaining genera, recognizable as the sole Onagraceous group that lacks a floral tube extending as a floral tube from the tip of the ovary to the point where sepals and petals spread out. Illustrations below, borrowed from Wagner, et al, 2007, help interpret that character.







A few years ago, we’d have encountered a more complex family, but people have busily reinterpreted the generic limits of Onagraceae, to the point that we consider every plant in the Florida flora as one of two genera, Oenothera or Ludwigia. Of course that means we face sixteen species in the first and twenty-nine in Ludwigia, so the task of identification is unchanged – it just appears simpler. And if we were dealing with a West coast flora, there’d be more complexity, in that the center of diversity for Onagraceae is on the other side of the Rockies.]
For any of us who are older, the shift makes keeping track a bit more complex. I was comfortable with the upright Southern Bee Balm as a Gaura, Gaura angustifolia specifically, with its narrow leaves. But it can’t be; molecular data suggest the genus, which was on the botanical ‘Mayflower’ (i.e. Linnaeus’s 1753 Species Plantarum) with Oenothera shouldn’t be regarded as a separate genus. That means, nomenclaturally, there’s a winner and a loser; Gaura lost and Southern Bee Balm (described as G. angustifolia in 1803, by Michaux) suddenly becomes Oenothera simulans, falling back on a segregate name Small had published in 1905.
So how did this happen? In 2007, Wagner, Hoch, and Raven published an extensive review of the entire family, consolidating decades of research; Wagner maintains a website on the family also. Extensive effort was dedicated to explaining the expanded generic limits of Oenothera, which is “strongly supported by molecular data, … marked by at least two morphologicdal synapomorphies; the presence of an indusium on the style, and a lobed or peltate stigma,”

Terminology Useful in Studying Onagraceae:
- Caducous – abscissing. or shed, at maturity, often said of stipules in Onagraceae
- Vespertine – referencing flowers that open in the evening, at Vespers
- Floral tube – the hypanthium (a tubular combination of sepal and petal tissue that lifts the apparent sepals and petals above the inferior ovary in most Onagraceae, lacking in Ludwigia (and the key distinguishing character)
- Valvate – describing sepals that develop without overlapping, simply touching or sealed along their margins (see photo of Ludwigia)
- Indusium – a cup-like expansion of the style, just below the point of , bascially embracing, the branching or spreading stigmatic lobes in many Onagraceae – present in Oenothera but not as evident as in more tropical genera.
- Peltate – disk-like or shield-shaped, terminating a stalk, said of stigmas for certain Oenotheras. Think of the leaf -shape of Lotus or Hydrocotyle
- Commissural stigmas – stigmas in which the branching or lobing reflects lateral stigmas commensurate to each separate carpel (Brown, 1843, who suggested each carpel, primitively, produced two lateral stigmas)
- Ektexine (Ectexine, Sexine) – The structural outer layer of a pollen grain external coating (the exine)
- Paracrystalline beaded pollen ektexine – “the unique “paracrystalline” ektexine is one of the features that distinguishes the family from other core families of Myrtales” (Daghlian, et al, 1984)
- Palynomorph – a microscopic, organic-walled microfossil (in this case, pollen grains of Oenothera) that retain significant structural detail (thus are resistant to decomposition) such as to remain identifiable, giving clues to past environments and ecosystems.
- PTH (Permanent Translocation Heterozygosity) – From Johnson and Godfrey, 2024: “Many plant species in the genus Oenothera have a genetic system called permanent translocation heterozygosity (PTH), which renders plants functionally asexual when they self-fertilize. PTH results from reciprocal translocations of chromosomes that suppress recombination and a balanced lethal mortality of homozygous gametes that prevents independent assortment of alleles. When PTH plants self-fertilize, the offspring are genetically identical to the parents, but when they outcross they can create new genotypes. “
- Comose – bearing trichomes, describing seed of Tribe Epilobieae (not in our flora)
- EPO (Evening Primrose Oil) – seed oil of Oenothera which contains gamma linoleic acid, suggested as a potential treatment for eczema
- Synapomorphy – one of many important terms used in Cladistic discussion: syn = combining; apo = to separate; morphy = form or shape: “a characteristic present in an ancestral species and shared exclusively with the evolutionary descendants” Oxford Languages. In this case, the pollen outer wall (ektexine, or ectexine) of many Onagraceae, described as “polycrystalline beaded” in texture, is a character that segregates Onagraceae from other Myrtales.
The CARYOPHYLLS & SUPERASTERIDS
Here we are really speaking of two separate branches. The Caryophylls have long been recognized as distinct, often set aside from other dicots. The most recent suggestion from the APG has been to anchor them somewhere at the base of the Asterids. As a group, they are preponderantly herbaceous, which means many of Florida’s Wildflowers are to be encountered in these several families. Following the outline of Families and Genera given below, we need to construct a basic working description of their nature.
The other two orders, Cornales and Ericales, are considered more closely related to the two main branches of Asterids (the Lamiids and Commelinids). The Cornales are mostly woody plants, but the Ericales include many herbaceous groups. The Ericaceae includes some curious herbaceous representatives, and our Wildflower flora boasts a few native Polemoniaceae (Phlox) as well as Primulaceae (Lysimachia). But the most celebrated native Wildflowers tucked in the Ericales are Sarraceniaceae, the Pitcher Plants. After discussing significant Caryophylls, we’ll certainly dip into the Sarracenia for a bit of technical detail.
- Caryophyllales: Cactaceae (Native 6/14 Acanthocereus, Consolea, Harrisia, Opuntia, Pilosocereus, Rhipsalis; Exotic 6/11 Cereus, Epiphyllum, Opuntia, Pereskia, Rhipsalis, Selenicereus), Caryophyllaceae (Native 9/23 Arenaria, Silene, Stellaria, Geocarpon, Paronychia, Sabulina, Sagina, Spergularia, Stipulicida; Exotic 15/24 Agrostemma, Arenaria, Atocion, Cerastium, Dianthus, Drymaria, Gypsophila, Polycarpaea, Polycarpon, Saponaria, Scleranthus, Silene, Spergula, Stellaria, Vaccaria), Droseraceae (Native 1/5 Drosera; Exotic 1/1 Dionaea), Nyctaginaceae (Native 4/7 Boerhavia, Guapira, Okenia, Pisonia; Exotic 2/2 Boerhavia, Mirabilis:, Petiveriaceae (Native 2/2 Petiveria, Rivinia) Phytolaccaceae (Native 2/2 Phytolacca, Trichostigma), Plumbaginaceae (Native 2/2 Limonium, Plumbago: Exotic 1/1 Plumbago), Polygonaceae (Native 7/29 Brunnichia, Coccoloba, Eriogonum, Fallopia, Persicaria, Polygonum, Rumex; Exotic 9/22 Antigonon, Emex, Fagopyrum, Fallopia, Persicaria, Polygonum, Reynoutria, Rumex:, Portulacaceae (Native 1/2 Portulaca; Exotic 1/4 Portulaca)
- Cornales: Cornaceae (Native 1/5 Cornus), Hydrangeaceae (Native 2/4 Hydrangea, Philadelphus; Exotic 1/1 Deutzia), Loasaceae (Native 1/1 Mentzelia), Nyssaceae (Native 1/4 Nyssa)
- Ericales: Balsaminaceae (Native 1/1 Impatiens; Exotic 1/2 Impatiens), Clethraceae (Native 1/1 Clethra), Cyrillaceae (Native 2/2 Cliftonia, Cyrilla), Ebenaceae (Native 1/1 Diospyros; Exotic 1/2 Diospyros), Ericaceae (Native 17/33 Agarista, Bejaria, Ceratiola Chimaphila, Epigaea, Eubotrys, Gaylussacia, Hypopitys, Kalmia, Leucothoe, Lyonia, Monotropa, Monotropopsis, Oxydendrum Pieris, Rhododendron, Vaccinium), Polemoniaceae (Native 2/7 Ipomopsis, Phlox; Exotic 1/1 Phlox), Primulaceae incl. Myrsinaceae, Samolaceae & Theophrastaceae (Native 5/9 Ardisia, Jacquinia, Lysimachia, Myrsine, Primula; Exotic 4/7 Ardisia, Bonellia, Jacquinia, Lysimachia), Sapotaceae (Native 3/13 Chrysophyllum, Manilkara, Sideroxylon; Exotic 3/4 Manilkara, Mimusops, Pouteria Sarraceniaceae (Native 1/7 plus 9 named hybrids Sarracenia) Styracaceae (Native 2/4 Halesia, Styrax), Symplocaceae (Native 1/1 Symplocos), Theaceae (Native 2/2 Gordonia, Stewartia; Exotic 1/1 Sphaeropteris).
The LAMIIDS
- Boraginales: Boraginaceae (Native 9/21 Andersonglossum, Bourreria, Euploca, Heliotropium, Lithospermum, Myosotis, Myriopus, Nemophila, Varronia; Exotic 9/15 Buglossoides, Cordia, Cynoglossum, Ehretia, Euploca, Heliotropium, Nama, Phacelia, Varronia)
- Gentianales: Apocynaceae (Native 17/42 Amsonia, Angadenia, Apocynum, Asclepias, Chthmalia, Cynanchum, Echites, Funastrum, Gonolobus, Matelea, Metastelma, Orthosia, Pattalias, Pentalinon, Rhabdadenia, Thysanthella, Vallesia; Exotic 18/ 24 Allamanda, Alstonia, Araujia, Asclepias, Calotropis, Carissa, Catharanthus, Cryptostegia, Gonolobus, Leptadenia, Nerium, Ochrosia, Plumeria, Rauvolfia, Tabernaemontana, Thevitia, Trachelospermum, Vinca), Gelsemiaceae (Native 1/2 Gelsemium), Gentianaceae (Native 6/22 Bartonia, Eustoma, Gentiana, Obolaria, Sabatia, Voyria; Exotic 1/1 Centaurium), Loganiaceae (Native 2/7 Mitreola, Spigelia; Exotic 1/1 Strychnos) Rubiaceae (Native 24/45 Borreria, Casasia, Catesbaea, Cephalanthus, Chiococca, Diodia, Edrastima, Erithalis, Ernodea, Exostema, Galium, Guettarda, Hamelia, Hexasepalum, Houstonia, Mitchella, Morinda, Oldenlandia, Pentodon, Pinckneya, Psychotria, Randia, Spermacoce, Strumpfia; Exotic 14/22 Borreria, Coccocypselum, Ixora, Mitracarpus, Morinda, Nauclea, Oldenlandia, Oldenlandiopsis, Paederia, Pentas, Psychotria, Richardia, Sherardia, Spermacoce
- Lamiales: Acanthaceae (Native 9/21 Avicennia, Dicliptera, Dyschoriste, Elytraria, Hygrophila, Justicia, Ruellia, Stenandrium, Yeatesia; Exotic: 14/31 Asystasia, Barleria, Carlowrightia, Eranthemum, Graptophyllum, Hygrophila, Hypoestes, Justicia, Nelsonia, Odontonema, Pseuderanthemum, Ruellia, Strobilanthes, Thunbergia, Bignoniaceae (Native 4/4 Amphitecna Bignonia, Campsis, Catalpa; Exotic 14/15 Amphilphium, Bignonia, Crescentia, Dolichandra, Handroanthus, Jacaranda, Newbouldia, Podranea, Pyrostegia, Radermachera, Spathodea, Tabebuia, Tecoma, Tecomaria), Lamiaceae (Native 21/69 Calamintha, Callicarpa, Clinopodium, Collinsonia, Conradina, Dicerandra, Hedeoma, Hyptis, Lycopus, Macbridea, Monarda, Ocimum, Physostegia, Piloblephis, Pycnanthemum, Salvia, Scutellaria, Stachydeoma, Stachys, Teucrium, Trichostema; Exotic 27/49 Ajuga, Cantinoa, Clerodendrum, Clinopodium, Coleus, Condea, Cornutia, Glechoma, Gmelina, Holmskioldia, Hyptis, Lamium, Leonotis, Mentha, Mesosphaerum, Monarda, Ocimum, Perilla, Petita, Pyrostegia, Premna, Prunella, Salvia, Scutellaria, Stachys, Vitex, Volkameria Lentibulariaceae (Native 2/20 Pinguicula, Utricularia), Linderniaceae (Native 2/5 Lindernia, Micranthemum; Exotic 2/3 Bonnaya, Torenia), Oleaceae (Native 4/11 Cartrema, Chionanthus, Forestiera, Fraxinus; Exotic 3/13 Jasminum, Ligustrum, Noronhia, Orobanchaceae (Native 10/29 Agalinus, Aphyllon, Aureolaria, Buchnera, Conopholis, Epifagus, Macranthera, Pedicularis, Schwalbea, Seymeria; Exotic 4/4 Aphyllon, Castilleja, Orobanche, Striga), Phrymaceae (Native 1/1 Mimulus, Phryma, Plantaginaceae (Native 11/33 Bacopa, Callitriche, Gratiola, Linaria, Mecardonia, Penstemon, Plantago, Scoparia, Sohronanthe, Veronica, Veronicastrum; Exotic 14/25 Angelonia, Bacopa, Callitriche, Kickxia, Leucospora, Limnophila, Linaria, Maurandya, Misopates, Plantago, Russelia, Scoparia, Stemodia, Veronica), Scrophulariaceae (Native 2/2 Capraria, Scrophularia; Exotic 3/7 Bontia, Buddleja, Verbascum), Tetrachondraceae (Native 1/1 Polypremum), Verbenaceae (Native 7/21 Citharexylum, Gladularia, Lantana, Lippia, Phyla, Stachytarpheta,Verbena; Exotic 8/17 including 4 hybrids, Duranta, Gladularia, Lantana, Lippia, Petrea, Phyla Stachytarpheta, Verbena)
- Solanales: Convolvulaceae (Native 9/43, Calystegia, Cuscuta, Dichondra, Distiake, Evolvulus, Ipomoea, Jacquemontia, Stylisma; Exotic 10/27 Aniseia, Camonea, Convolvulus, Cuscuta, Dichondra, Distimake, Evolvulus, Ipomoea, Jacquemontia, Poranopsis), Hydroleaceae (Native 1/3 Hydrolea; Exotic 1/1 Hydrolea),, Solanaceae (Native 5/22 Capsicum, Datura, Lycium, Physalis, Solanum; Exotic 12/39 Browallia, Brugmansia, Calibrachoa, Cestrum, Datura, Hyoscyamus, Nicandra Nicotiana, Petunia, Salpichroa, Solandra, Solanum
LAMIALES
LAMIACEAE
Studying Judd, et al (2016) you’ll feel a bit unsettled as to the delimitation of the sixteen families currently recognized in the Order called Lamiales. There’s solid familiarity here, with the Olives, Foxgloves, Catalpas, Lantanas, and Mints, a group of plants that mostly show flowers with few stamens (normally 2 or 4) and tubular, zygomorphic (bilaterally symmetrical) corollas, along with a tendency to bear opposite leaves. The families fall out separately based on a variety of characters, generating taxonomic turmoil over the last three decades.
Two families (as well as certain genera, i.e. Scoparia) in the Order stand out as distinctive (Oleaceae and Tetrachondraceae, i.e. Polypremum) in having radially-symmetrical flowers. Following determinations that the Scrophulariaceae was a mixed bag, a much-tighter family was defined, with Orobanchaceae and Plantaginaceae inheriting the misfits. The bulk of families, including the Lentibulariaceae (Bladderworts), Bignoniaceae (Cross-vine), Acanthaceae (Ruellia), Verbenaceae (Lantana), and Lamiaceae are those with zygomorphic flowers in a 2-3 lobing pattern (an upper lip of 2 lobes and lower lip of 3 lobes). There are incongruent determinations, such as the grouping of Scoparia with Antirrhinum (Snapdragon), Digitalis (Foxglove), and Plantago (Plantain) in the Plantaginaceae. Make it make sense. As we say in the South, “somethin’ ain’t rite.”


Judd et al tell us differences among Lamiaceae, Verbenaceae, Acdanthaceae, Bignoniaceae, are slight and we might well learn those and other currently recognized families belong together as one. Stay tuned.
Meanwhile, the Mints (Lamiaceae, also Labiateae) as currently defined, with 250+ genera and over 7100 species, are an extensive worldwide plant family, particularly rich in herbaceous and suffrutescent representatives. Even given the close relationship to other plant families, the Mints are viewed as a related assemblage of of common origin (monophyletic). It’s also a reasonably-easy group for field identification, recognized by many shared characters – opposite, decussate leaves (often with square stems) that are often distinctly fragrant with essential oils, flowers of 5 lobes, forming an upper lip of two and a lower lip of three lobes.

There are either two or four stamens, paralleling an elongated style, asymmetrically-bifid with a small stigmatic surface.

Families most easily confused with Mints are the closely related Acanthaceae, Bignoniaceae, and Verbenaceae. The Verbenaceae are generally distinguished by their more rotate corollas (a narrow tube expanding to form a flattened, nearly rotate corolla). I pick out Bignoniaceae by their characteristic bow-legged filaments and fused anthers, as well as a tendency toward woodiness and compound leaves. The Acanths fall out, in the field, by a general presence of obvious leafy floral bracts or conspicuous sepals, and (for lack of a better description) their consistently cockeyed stamens.


Using data from the ISB Florida Plant Atlas, one encounters curious taxonomies. The flora lists 118 taxa distributed in 41 genera, suggesting many genera with modest representation, which is what we discover. Twenty genera are represented by a single species. That coincides with the observation that we also see twenty genera of non-native Mints, but they are not the same twenty genera. Thirteen non-native Mint genera are represented by a single species, and another five by two species. Only the exotic Vitex (5 species) and Clerodendron (9 species) are more diverse.

Of native Mints, we find there are twenty-two genera (sixty-nine species and varieties). Eight genera include both native and non-native plants, for example Monarda, Physostegia, Salvia, and Scutellaria, while the other fourteen include no exotics. Of our 69 native Mint taxa, seventeen (in 5 genera) are listed as endemic, as can be seen in the spreadsheet below:

Sort of fascinating…. I’m certain these many segregates are well-studied and well-documented, but one has to wonder if this level of discrimination relates to geographical history and plant biology, or is actually an artifact of relatively more dedicated field study. Plant taxonomy is something of an individual pursuit, and one of the lesser-funded sciences. Our understanding of different plant groups is patchy, some are easier to study, or have attracted the attention of entire groups of scholars. Mints call attention to themselves and make great herbarium specimens, while other kinds plants may be less common, or less easy to voucher. For some reason, it seems variation and geography of Mints have been either better documented or more intensely examined than in other groups of plants. But that’s a very superficial observation.
SOLANALES
CONVOLVULACEAE

It’s worth a moment to simply get our heads around the Convolvulaceae. The family name is typified by Europe’s notorious Field Bindweed, Convolvulus arvensis (convolvulus = entwining; arvensis = of fields and pastures), an invasive plant that frequents ruderal areas and plagues North America’s farmers and home gardeners, but is said to be less problematic in native habitats.

Field Bindweed is reported only from very western counties in Florida’s Panhandle, but probably covers far more area. I like this statement from Wikipedia: “Bindweed can be controlled by pulling it out by hand or plowing it up every three weeks, for three up to seven years.“
The family includes one significant crop, Ipomoea batatas, variously called Sweet Potato, Batata, even Yam (an unfortunate and confusing use). Native to Pacific coastal areas of the New World tropics, Sweet Potato is a frost-intolerant vine introduced and widely cultivated throughout tropical and warm-climate areas as a significant food source. Curiously, Europeans knew “batata” as potato before Solanum tuberosum (the other potato) was introduced from South America. It was the Sweet Potato (thought aphrodisiac) Falstaff petitions in ‘The Merry Wives of Windsor’ with the phrase “Let the sky rain potatoes.”

Yes, these are Morning-glories, but there’s a bit more to the story. The Florida Plant Atlas shows thirteen genera, four of which are non-natives represented by a single species each (Anisela, Camonea, Convolvulus, and Poranopsis), three small genera show only native representation (Bonamia, Calystegia, and Stylisma), and the bulk of species in our flora are genera with both native and non-native representation (Cuscuta, Dichondra, Distimake, Evolvulus, Ipomoea, and Jacquemontia).
Two genera should be called out (Cuscuta and Dichondra) as so distinct from the others as to sometimes segregate in their own families. The Dodders (Cuscuta spp) are distinctive in many regards, the most obvious point being their achlorophyllous parasitic lifestyles. Most Dodder species in our flora are native, but the more commonly encountered is an invasive exotic, Cuscuta japonica. You might question assignment of Cuscuta to the Convolulus family, but they are vines, and at least are readily identified; they have to go somewhere. Dichondra is another plant questionably assigned to this family, with its distinctive ovary (the style forms from the base). But given their trailing habit, small leaves, and diminished corollas (about equal to or shorter than the calyx), the Dichondras will hardly vex us.
Of seventy taxa reported in the Atlas, over forty are considered native, and most of those fit the family mold. That means their short-lived flowers form conspicuous, tubular funnel-shaped corollas varying from white to various anthocyanin-rich blues, purples, and pinks. Once you’re convinced a plant identifies as Convolvulaceous, three genera (Bonamia, Evolvulus, and Stylisma) can be quickly sorted out as having two styles attached to the ovary apex, which leaves seven genera to worry about. Three of those are tropical non-natives (Aniseia, Camonea, and Poranopsis), collected only in the southern-most counties, which means for the most of Florida, there are only four genera to puzzle out – Jacquemontia, Calistegia, Ipomoea, and the noxious bindweed Convolvulus. Of thirty-six taxa in these four genera, twenty-nine are Ipomoea, which means the main taxonomic task for Convolvulaceae is deciphering that genus.

Our native Ipomoea sagittata has a history of reverse fortune. Native to marshes and saltwater edges along the Gulf coast and into the Caribbean, the plant also grows along the coast of North Africa, where it was encountered and described by Jean-Louis-Marie Poiret (a botanist and explorer sent by the French King to explore Algeria) in the 2nd volume of his Voyages en Barbarie…., 1789.

Terms Useful in Studying Convolvulaceae
Bine – a word describing vines that climb strictly through helical (twining) growth (without tendrils or clasping elements), hence the term bindweed used for Convolulus and other plants in this family
Camote – a common term for Sweet Potato in Latin America
Slips – propagation stem cuttings taken from stored tuberous roots
Amukeke, Inginyo – dried Sweet Potato root products important in Africa
Cymose – a general term for the short inflorescence we see in Ipomoea, with a terminal flower, flanked by a few axillary flowering branches – sometimes encountered in unreviewed internet publications as “zymous”
GENTIANALES
APOCYNACEAE

Students in the last Century were accustomed to learning the Dogbanes (Apocynaceae) and Asclepiads (Asclepiadaceae) were paired families. Both produced copious white latex, both tended toward opposite leaves, and both produced fruit in simple pods that were often paired. The Apocynaceae (Dogbane, Oleander, and a host of tropical woody plants) produced somewhat tubular petaloid flowers with curiously compact, united , barely visible sexual parts; Asclepiads were predominantly herbaceous, and bore unmistakably characteristic flowers geared to meticulous pollination mechanisms. Whether native to North America, the Mediterranean, or Africa, there’s little mistaking an Asclepiad from other plants.
Contemporary analytical methods convince systematists that Asclepiads, though clearly distinct, form a branch surrounded by plants we’ve considered Apocynaceae, which means the Asclepiads have been “subsumed” – now forming a subfamily in Apocynaceae. Melding the two families recognizes the nearly indecipherably broad range of modifications characterizing flowers of many genera worldwide, from the most normal seeming to the unbelievably complex, that is from Apocynum to the Asclepius, a claim easily stated and nearly impossible to explain in simple terms. If you don’t want to wade into that swamp, follow this link straight to Asclepias. Otherwise, stay the course
Many features unite the group – general presence of lacticifers producing milky sap, 5-merous flowers (5 sepals, 5 petals, 5 anthers, 5 stigmas but 2 carpels), petals basally united to the stamens (often forming a ring or torus), an androecium, built of stamen filaments and 2-loculate anthers capable of evolving remarkable structural detail, with pollen loosely aggregated (by viscin strands) varying to fused and solid, and a gynoecium of two separate carpels curiously united by a capitate “stylar head”, the stigmas positioned laterally or imbedded in chambers between the anthers. In the more complex floral structures, the androecium (combined staminal tissues) and gynoecium (combined pistillate tissues) create a miniature wonder called the “gynostegium” – a 5-sided pollination contraption generating nectar in stigmatic chambers and/or cuculli (we’ll get to this).
Simpler floral formats in the family include many common non-natives, Catharanthus for example. Below is an illustration borrowed from Rosatti (1989) and Judd et al (2019), detailing that floral structure. Note various issues: 1. In the mature bud (far left) you see the swollen, bulbous zone below the twisted petal lobes. That’s where the androecium (staminal tissue) develops, shown in long section in the flower dissection (center). 2. The style and stigma form a thumbtack-looking presence just below the point of attachment for the anthers, the top is the sterile “stylar head,” just below which is the hour-glass shaped stigmatic zone. 3. The two free carpels (individual ovaries) form on the receptacle, at the base of the corolla tube, joining at their tips as a single style. Alternating with the carpels are two nectaries (shown in the two figures bottom right).



Catharanthus provides a good, commonly-available introduction to the floral structure of the native Amsonia. Once again, Rosatti comes to our rescue with excellent diagrams to compare against material you may encounter in the field. Note anthers with short filaments attached inside the corolla tube, hovering above a stylar head, as in Catharanthus. In the top center, you see developing carpels (h) that become long narrow follicles (figure i., far left).

Robert Wyatt and Sara Lipon (2007) diagram and explain evolutionary steps that led to fusion of pollen (into pollinia) as well as loss of carpel fusion (such that pollen does not hold free reign to fertilize either of the two carpels.) They showcase this with four examples, beginning with the North Temperate Apocynum (A & B) which has a stylar head that combines pollen tube access to the two carpels through part of the style called a “compitum”, moving then through two Afro-Asian plants, Periploca (C & D) in which anthers unite to form a column around the style, and Secomone (E & F), which shows formation of a complex gynostegium, with pollen massing in obvious sacs, while pollen tube access to both carpels remains through the “compitum.” Their discussion moves, finally to Asclepias (G-J), which shows solid pollinial masses and development of stigmatic chambers, while sacrificing functions of the compitum. (see their illustrations below.)




Study of the many morphological developments shown in illustrations above helps explain the reasons the Asclepiads have been subsumed to the Apocynaceae. The more limited selection of plants in our flora scrubs the Apocynaceae (with 400 genera and over 4500 species) of intermediate stages, leaving an impression of clear distinctions that doesn’t bear out when studying these plant worldwide.
We find ISB lists thirty-three genera of Apocynaceae for Florida, which fit into four subfamilies – 1) Subfamily Rauvolfioideae: Native: Amsonia, Vallesia. Non-native: Allamanda, Catharanthus, Carissa, Ochrosia, Plumeria, Rauvolfia, Tabernaemontana, Thevetia, Vinca. 2) Subfamily Apocynoideae: Native: Angadenia, Apocynum, Echites, Pentalinon, Rhabdadenia; Thyrsanthella. Non-Native: Nerium, Leptadenia, Trachelospermum. 3) Sub-family Periplocoideae: Non-native: Cryptostegia, Alstonia. 4) Sub-family Asclepiadoideae: Native: Asclepias, Chthamalia, Cynanchum, Funastrum, Orthosia, Pattalias, Metastelma, Matelea. Non-native: Araujia, Calotropis, and also non-natve Asclepias and Gonolobus.
Fifteen genera (20 species) include no native species; almost all are of the subfamilies with tubular flowers, twisted petals, and unconsolidated gynostegia. Native Apocynaceae are represented by sixteen genera, two of which (Asclepias and Gonolobus) include one exotic species. Several are of the simpler Apocynoid type (Apocynum, Amsonia, Angadenia, Echites, Pentalinon, Rhabdadenia, and the odd-man-out Thyrsanthella), while most are in the Asclepiad sub-family (Asclepiadoideae). We find the genus Asclepias accounts for twenty-one native taxa. In summary, the family includes 33 genera, almost all of which include only one or two species. The native Matelea (4 species) and Asclepias (22 species) stand out.
That brings us to Asclepias, which, as the most species-rich genus in our regional flora merits close attention. To appreciate the minute and complex structure (and functionality) of these flowers, we have to begin with whichever milkweed you know already. Woodson’s significant monograph (1954) remains valid, even though published 70 years ago. He treated all 22 species included in the Florida Plant Atlas, including distribution maps and a simple line drawing for each taxon. Using Photoshop, I lifted and cleaned his line drawings (“beautifully illustrated by Mrs. Froeschner and Mr. Heinze”) from an original copy of the text.






















They all share the same unique flower structure, which is 5-merous (5 petals, 5 sepals, 5 anthers, and 2 carpels with 5 stigmatic chambers). In most (not all) species, the petals are reflexed (turned downward when flowers open), exposing a crown-like corona, made of 5 “cuculli”, each constituting a scoop-like “hood” surrounding a tongue-like projection called the “horn”.



In the central axis, surrounded by the corona, sits a very complex structure, the “gynostegium,” combining and positioning anthers and stigmas. Pollen in the five anthers coalesces, unbelievably, into ten pollinia (one pollinium in each of two anther chambers) that look like microscopic tennis rackets. The head of each racket is of melded pollen grains, while the grip (called a translator arm) ends at a sticky pad (the corpusculum, or gland) that can attach to the leg (tarsus) of the right visiting insect. In fact, the same pad, which is positioned between two anthers will serve two pollinia (one from each flanking anther). If successfully removed, the next incredible trick involves a racket-like pollinium slipping into a slit in that or another flower (NOTE: Most Asclepias have been shown to be self-incompatible (infertile when self-pollinated), though there are suggestions the level of compatibility might vary between populations)
Yes, it’s complex, and yes, Asclepiads are known for low fruit yield. But each fruit yields many wind-borne seed. Somehow, it works. Asclepiads, though seldom found in dense stands manage to get around, establish, and procreate. And there are many species, over 200 for the genus, 21 of which are native to Florida.
Quoting directly from Bob Woodson, in his extensive 1941 Perspective of the Genera (of North American Asclepiads): “The method of pollination in Asclepiads which Robert Brown disclosed appears at first sight to be of design so cunning that great fecundity of the plants should be assured, as well as frequent hybridization. Every conceivable innovation would seem to have been made by the flowers to insure successful insect ministrations. But such does not necessarily seem to have resulted. The family as a whole is outstanding for the constancy of its species and the rarity of obvious hybridity. Furthermore, the amount of fruit produced scarcely seems to be commensurate with the ‘‘efforts’’ taken to insure it, to speak anthropocentrically.”
Plants in the Dogbane side of the family (the two sub-families Apocynoideae and Rauvolfioideae), at least in aspect, are so very different. Many you’ll meet in Florida are non-native, common horticultural introductions, such as Oleander (Nerium), Star Jasmine (Trachelospermum), Periwinkle (Vinca and Catharanthus), Natal Plum (Carissa), and Allamanda. These are plants valued for their rich green, leathery foliage and showy flowers, generally producing campanulate or salverform corollas noted for the sculptural swirl petals inherit from the twisted shape characterizing developing buds. Those features, to a lesser extent, characterize the native Apocynum, Amsonia, etc.
For Florida field botanists, the distinction is significant. Recognizing a plant as an Asclepiad and recognizing the rest of the Apocynaceae require internalizing two distinct gestalts. [See the International Asclepiad Society]
Terminology Useful when studying Apocynaceae
- Corona – a crown of anther tissue surrounding the gynostegium, formed by five cuculli – each cucullus being a specialized outgrowth from a stamen, consisting of a hood (a nectary) which usually will surround a projection called a horn.
- Faucal Annulus – a ring of tissue formed from the corolla (complexed around the corona), also referred to as the coralline annulus. Frye suggests the corolla of Aclepiads emerges from a torus, which would poit to formation of the annulus
- Gynostegium – a complex unified structure that combines the anthers with their accessory enations and sequestered stigmatic cavities, each accessed through a slit. The five (typically) stigmatic cavities actually correspond to only two carpels.
- Compitum – a shared transmiting track in many Apocynaceae that allows pollen deposited on one stigma to fertilize either carpel, absent in Asclepias (Wyatt & Lipow, 2007),
- Stigma head – the somewhat flattened top of the gynostegium, also termed the style table, which Frye (1902) explains results from fusion of the tips of the two carpels.
- Staminal wings – lateral projections from either side of both anther chambers, forming guide rails that direct pollinia to the alar slit, the entrance to the stigmatic chamber. This entry is directly below the corpusculum
- Filament tube – the column, a tube formed by union of stamen filaments
- Pollinium – the aggregated pollen in each chamber of an anther
- Viscin –
- Translator arm – a stiff stipe, i.e. arm that attaches a pollinium to a corpusculum (gland). The translator is not cellular, rather a hardened resin secreted by glandular tissue into a hollowed channel that functions as a mold.
- Translator apparatus – the translator arms (each of which carries a pollinium) from locules of two adjacent anthers and the attached corpusculum
- Pollinarium – the pair of pollinia, each from a separate anther, as wee as the entire translator mechanism
- Caudicle – the portion of the translator arm that attaches directly to the pollinium. The portion of a translator arm that remains once a pollinium is detached is called the retinaculum
- Corpusculum – also called a gland, a somewhat folded, sticky pad (secreted by a gland) that is pried open and then closes onto/attaches to insect visitors, carrying two translator arms
- Pappus – the crown of single-celled hairs (formed from epidermal cells near the micropyle) that develops as an aide in seed dispersal (Frye, 1902)
- APSA clade – referencing four subfamilies, the Apocynoideae, Periplocoideae, Secamonoideae, an Asclepiodoideae (Wyatt & Lipon, 2007)
The CAMPANULIDS
- Apiales: Apiaceae (Native 18/36 Angelica, Centella, Chaerophyllum, Cicuta, Cryptotaenia, Daucus, Eryngium, Ligusticum, Lilaeopsis, Oxypolis, Ptilimnium, Sanicula, Sium, Spermolepis, Thaspium, Tiedemannia, Trepocarpus, Zizia; Exotic 13/15, Ammi, Anethum, Apium, Bowlesia, Bupleurum Coriandrum, Cyclospermum, Daucus, Eryngium, Foeniculum, Petroselinum, Scandix, Torilis), Araliaceae (Native 2/5 Aralia, Hydrocotyle; Exotic 6/9 Eleutherococcus, Hedera, Heptapleurum, Hydrocotyle, Polyscias, Tetrapanax), Pittosporaceae (Exotic 1/2 Pittosporum)
- Aquifoliales: Aquifoliaceae (Native 1/13 Ilex; Exotic 1/1 Ilex)
- Dipsacales: Caprifoliaceae (Native 4/4 Lonicera, Symphoricarpos, Valeriana, Valerianella; Exotic 2/2 Abelia, Lonicera), Viburnaceae incl. Adoxaceae 2/6 Sambucus, Viburnum; Exotic 1/1 Viburnum)
- Asterales: ) Campanulaceae (Native 3/20 Campanula, Lobelia, Triodanis; Exotic 2/3 Hippobroma, Wahlenbergia), Menyanthaceae (Native 1/2 Nymphoides; Exotic 1/4 Nymphoides), Asteraceae (Native 87/314; Exotic 74/ 107 – Genera Native only: Achillea, Ageratina, Ambrosia, Antennaria, Arnica, Arnoglossum, Baccharis, Balduina, Berlandiera, Bigelowia, Boltonia Borrichia, Brickellia, Brintonia, Carphephorus, Chaptalia, Chrysogonum, Chrysoma, Chrysopsis, Conoclinium, Croptilon, Doellingeria, Echinacea, Eclipta, Elepantopus, Erechtites, Eurybia, Eupatorium, Euthamia, Eutrochium, Fleishmannia, Garberia, Hartwrightia, Hasteola, Helenium, Heliopsis, Heterotheca, Hieracium, Hymenopappus, Ionactis, Koanophyllon, Krigia, Liatris, Lygodesmia, Marshallia, Melanthera, Nabalus, Oclemena, Packera, Phoebanthus, Pluchea, Pterocaulon, Pyrrhopappus, Rayjacksonia, Rudbeckia, Sachsia, Sclerolepis, Sericocarpus, Silphium, Smallanthus, Solidago, Stokesia, Tetragonotheca, Vernonia; Genera Mixed, Native and Exotic: Acmella, Ageratum, Bidens, Chromolaena, Cirsium, Coreopsis, Erigeron, Flaveria, Gaillardia, Gamochaeta, Helianthus, Hypochaeris, Iva, Lactuca, Mikania, Palafoxia, Pectis, Pityopsis, Pseudognaphalium, Ratibida, Symphyotrichum, Verbesina; Genera All Exotic: Acanthospermum, Anthemis, Aphanostephus, Artemisia, Bradburia, Calyptocarpus, Centaurea, Centratherum, Cichorium, Cladanthus, Cosmos, Crassocephalum, Crepis, Cyanthillium, Dittrichia, Emilia, Enydra, Euryops, Facelis, Filago, Galinsoga, Gerbera, Glebionis, Gynura, Haplocarpha, Lagascea, Launaea, Leucanthemum, Melampodium, Onopordum, Parthenium, Pascalia, Pilosella, Polymnia, Praxelis, Pseudelephantopus, Pseudogynoxys, Pulicaria, Senecio, Soliva, Sonchus, Sphagneticola, Synedrella, Tagetes, Taraxacum, Tithonia, Tridax, Xanthium, Youngia, Zinnia
ASTERALES
asteraceae
We dive into the DAISIES – the Composites, the Asters, the Asteraceae – as a familiar group. Almost anyone has had some encounter with a Daisy, a Sunflower perhaps. There’s hardly a more iconic plant.

Native to much of western North America, Helianthus annuus has gotten around, showing up along roadsides throughout Eastern North America and as edible seed in snack counters worldwide. Sunflower seed were among the earliest New World treasures to gain a foothold in Europe, with productive strains selected by Russian agronomists for cooking oil production (see Daniil Bokarev, who is credited with developing industrial scale methods for extracting oil).
In Florida, you’ll run into Helianthus annuus and another non-native, H. tuberosa (Jerusalem artichoke), along with 16 native Helianthus species, such as the compact H. debilis and the curious H. radula – which is classified as Helianthus for reasons I’ve yet to explore.


Iconic as sunflowers may be, your daily encounter with members of the Daisy family (alias Composites, formally the Asteraceae) is likely more mundane. Perhaps at a salad bar you select one of the various strains of lettuce (Iceberg, Romaine, etc.), hundreds of varieties having been coaxed from the gene pool of Lactuca sativa beginning along the Nile by pyramid-constructing Egyptians.


Sunflower and lettuce, so very different in many ways, cling together with over 30,000 other species as a clearly distinct branch on the tree of life, identified readily by their flower and fruiting structures. We call them Composites and systematize them as the Asteraceae (also the Compositae) because they all generate very tight heads (capitulae) composed of small flowers (florets), each head of florets presenting itself as an individual flower, so thoroughly deceptive as to have earned the designation “pseudanthium”, i.e. false flower. Even before opening, the flower heads appear as flower buds (perhaps we should call them pseudobuds), though they’re actually much more complex. The green outer segments are not sepals, rather leafy bracts (also called phyllaries), forming a precise envelopment called the involucre. Those involucral bracts provide the first of several clues that a plant belongs to this family.





Of course, the prize for involucral bracts must be awarded to the Artichoke, a Mediterranean plant that, like lettuce, was selected for its tasty usefulness (though some might disagree as to how tasty they are.) Artichokes, with their exaggerated capitulae (heads) and choke of disk florets, remind us that thistles are Composites also.



Given a single floret, almost anyone familiar with these plants would know the floret came from a Composite. The first obvious character is the inferior ovary, a term that tells us the flower parts are seen atop the fruiting body (inferior = below), which in this case looks like a stout stalk. But you’ll search in vain for a calyx (the collective term for sepals), finding a short tube of petals that is surrounded at its juncture with the ovary by a “pappus” – a ring of bristles, teeth, or scales, or sometimes nothing at all. The pappus alone lets us know this is a Composite floret. Rising from inside the pappus, the tube formed by the petals will be one of two radically-distinct forms, either a trumpet-shaped funnel or a conspicuously lopsided deal variously called a “ray” or a “ligule” (some writers distinguish rays from ligules based on how completely the five petals participate in this 1-sided structure).

And, yes, there are intermediates – Check out the florets of Stokesia below.


That’s not all. Spotting Composite florets is a world of riches in that the anthers and stigma are as singular as other flower components. In fact, it’s the 5 anthers that give rise to a term for the study of composites – “synantherology” – the “syn” (which means combined or fused) reminding us that Composite anthers cling together, surrounding the emerging style and stigma like a collar or tube (called the “theca”). As if this were not enough to seal the deal, the final stage comes when the bifid style pushes up and out through the collar of anthers, peeling back in a forked or rolled pair, like a fountain making it’s show.
From their highly characteristic flowers, Composites mature fruit that are just as recognizable – always hard, 1-seeded structures that do not dehisce (they do not crack open naturally to release seed). You’ll see these referred to as “achenes” or “cypselae” – the term cypsela being restricted to Composites. This, of course, means that technically the black and white striped hull of a Sunflower “seed “is the fruit wall, not simply a seed coat, which also means you should feel a slight tang of botanical guilt when referring to them as “seed” – the seed is the loose thing you eat once it has been freed from the imprisioning fruit wall.
It’s common among Composites for the pappus to become more evident during fruit maturation, forming some functionally-important structure of dispersal, like the wind-catching parasols of Lygodesmia or the barbed anchors of Bidens.

Useful Terminology for Studying Composites (In this second link, check out Appendix A, Illustrated glossary of Composites, by Nádia Roque, David J. Keil, and Alfonso Susanna
- Synantherology – what specialists (synantherologists) in the study of Composites call their work
- Pseudanthium – a term that allows us to call the Composite inflorescence something other than a “flower” – which is a botanical point of angst
- Synflorescence – a yet broader term used when several heads (capitulae) of Composite flowers unify as a tertiary presence. Similar in meaning to Capitulescence – used in describing an an array of tightly-headed inflorescences, such as Composite heads (capitulae) produced in a regular inflorescence.
- Winged – when a flap of tissue forms and characterizes the edge of some botanical part, such as a stem or an achene, also termed alate when with fruit or seed, and decurrent when speaking of green leaf tissue coursing down a stem
- Capitulum – the technical term for a head of flowers in the Composites. Taking the form of a single flower, you’ll also see a capitulum described as a pseudanthium (see above).
- Radiate (vs. Radiant) head – the typical daisy capitulum, the center (the disk) filled with disk florets rimmed by petal-like ray florets. A Radiant head appears to bear disk and ray flowers, but the larger peripheral florets are modified disk flowers, as in Stokesia.
- Ligulate head – a capitulum composed of ligulate florets solely, as in Chicory or Lactuca. In some treatments (FloraQuest), Ligulate florets, specifically, are perfect, while imperfect florets with a Lamina would simply be called Rays.
- Discoid – a Composite captitulum composed solely of disk florets that are perfect, as in thistles and Liatris. Compare to Disciform, a head of disk (disc) flowers that each head bears a medley of florets that may be perfect or unisexual or neuter.
- Involucre – the characteristic arrangement of bracts (Phyllaries) that envelope a Composite capitulum, completely covering florets during development and remaining through maturity. Some Composites bear a subtending set of distinct bracts (Cosmos, Coreopsis, Bidens) termed a Calyculus.
- Receptacle – the stem on which flower parts develop, in Composites the broadened stem (surrounded by the involucred) on which the florets develop. The best part of an Artichoke
- Phyllary – technical term specific to the bracts of the involucre
- Floret (Disk, Ray, & Ligule) – the individual flowers that make up a Composite capitulum. Disk florets are tubular and mostly radially symmetrical. Ray florets develop petals asymmetrically, as a lamina (strap) to one side. In a pseudanthium, the rays are the elements that are plucked to predict the status of a love relationship. Ligulate florets are distinguished from Ray florets as being perfect, and showing all 5 lobes as teeth along the tip of the lamina.
- Syngenesious – the term used to remind us the five anthers of Composites develop adherent to one another. They are not quite connate (parts of similar origin when they grow together as one component), because they not so thoroughly melded.
- Theca – the tubular space formed around the style by the stamen filaments and anthers
- Pales/Palea – the more technical term for receptacular bracts (also called Chaff), typically subtending florets. They can be of many sorts, from scale-like to thin and insubstantial, to stiff and needling, to somewhat leafy. A receptacle lacking pales is termed Epaleate.
- Achene – a common and acceptable term for the 1-seeded, hard-walled fruit of Composites
- Cypsela – the term specific to Composites that denotes the one-seeded, hard-walled, inferior fruit (also often called an Achene) characteristic of the family.
- Pappus – the ring of bristles or scales at the top of the Composite fruit, sometimes multi-seriate, sometimes absent, often involved in dispersal.
REFERENCES & RESOURCES
Akulovy, Zoya. CalPhotos You will find this Internet Resource astonishing
Brown, Robert, 1843. On the relative position of the Divisions of Stigma and Parietal Placentae in the Compound Ovarium of Plants, The Annals and Magazine of Natural History….,XI: 35-42
Endress, Peter K. and Merran L. Matthews, 2012. Progrees and problems in assessment of flower morphology in higher-level systematics, Plant Systematics and Evolution 298: 257-276. https://www.researchgate.net/publication/257447274
Funk, Susanna V., Todd F. Stuessy, and R. J. Bayer, ed. Systematics, Biology, and Evolution of the Compositae. Download at: https://www.compositae.org/downloads/Systematics_evolution_and_Biogeography_of_Compositae.pdf
Hellström, Nils Petter, Gilles André, and Marc Philipe, 2017. Agustin Augier’s Botanical Tree: Transcripts and translations of two unknown sources, Huntia 16: 17-38
Johnson, Marc T. J. and Ryan M. Godrey, 2024 Outcrossing rates in the permanent translocation heterozygote Oenothera biennis L. (Onagraceae), Botany 102: https://doi.org/10.1139/cjb-2023-0067
Judd, Walter S., Christopher S. Campbell, Elizabeth A. Kellogg, Peter F. Stevens, and Michael J. Donoghue, 2016. Plant Systematics – A Phylogenetic Approach, Fourth Edition, Sinaner Associates, Inc, Sanderland, MA.
Kukkonen, Ilkka, 1984. On the inflorescence structure in the family Cyperaceae, Annales Botanici Fennici 21: 257-264 https://www.jstor.org/stable/23725865
Maarten J. M. Christenhusz and James W. Byng, 2016. The number of known plant species in the world and its annual increase, Phytotaxa 261:201-217. http://dx.doi.org/10.11646/phytotaxa.261.3.1
Stevens, P. F. (2001 onwards). Angiosperm Phylogeny Website. Version 14, July 2017 [and more or less continuously updated since]. http://www.mobot.org/MOBOT/research/APweb/.
Taylor, Walter Kingsley, 2009. A Guide to Florida Grasses (Digital), University of Florida Press
Vegetti, Abrelardo C., 2003. Synflorescence typology in Cyperaceae, Ann. Bot Fennici 40: 35-46
Wagner, Warren L., Peter C. Hoch, and Peter H. Raven, 2007. Revised Classification of the Onagraceae, Systematic Botany Monographs, v. 83. https://repository.si.edu/bitstream/handle/10088/7611/bot_Wagner_et_al_2007-Onagraceae-sm.pdf?sequence=1
Watson, L and M. J. Dallwitz, 1994 onward (2025). The Families of Flowering Plants: Descriptions, Illustrations, Identification, and Information Retrieval. A remarkable resource on plants.
Woodson, Robert E, 1954. The North American Species of Asclepias L., Annals of the Missouri Botanical Garden, 41: 1-211. https://www.biodiversitylibrary.org/page/16156875
Link to the Page: https://botanyincontext.com/all-in-the-family/
Link to PDF of Families in Hammer Guide: https://botanyincontext.com/families-treated-in-roger-hammers-guide/
Link to Flora of Florida Family Index: https://botanyincontext.com/flora-of-florida-family-index/
Editing & Formatting: Text Color Code to Family names: ac0930; Text Color Code to Order names: 487c44; Background Color for Major Groupings: 65ae69
Curiosa
Urena lobata (Malvaceae) – Wikipedia notes the invasive Ceasarweed (native to South America) is famous for it’s treatment and commentary in the secular 1751 Encyclopédie ou Dictionnaire raisonné des sciences, des arts et des métiers, in which the plant is treated succinctly (“Aguaxima, a plant growing in Brazil and on the islands of South America.”), such that Denis Diderot, one of the editors, felt compelled to follow the entry with a commentary, translated as follows: “Aguaxima, a plant growing in Brazil and on the islands of South America. This is all that we are told about it; and I would like to know for whom such descriptions are made. It cannot be for the natives of the countries concerned, who are likely to know more about the aguaxima than is contained in this description, and who do not need to learn that the aguaxima grows in their country. It is as if you said to a Frenchman that the pear tree is a tree that grows in France, in Germany, etc . It is not meant for us either, for what do we care that there is a tree in Brazil named aguaxima, if all we know about it is its name? What is the point of giving the name? It leaves the ignorant just as they were and teaches the rest of us nothing. If all the same I mention this plant here, along with several others that are described just as poorly, then it is out of consideration for certain readers who prefer to find nothing in a dictionary article or even to find something stupid than to find no article at all.”